

.
Bands result from membrane strip incubation.")
.
Bands result from membrane strip incubation.")






, diluted 1/600 was used as secondary antibody.")
 and mouse anti-beta tubulin Ab(#T0023) for 1 hour at 37°C. An AlexaFluor594 conjugated goat anti-rabbit IgG Ab(Red) and an AlexaFluor488 conjugated goat anti-mouse IgG Ab(Green) were used as the secondary antibody.
The nuclear counter stain is DAPI (blue).")
 by IF/ICC. The samples were fixed with PFA and permeabilized in 0.1% Triton X-100, then blocked in 10% serum for 45 minutes at 25°C. Samples were then incubated with primary Ab(#AF0863) and mouse anti-beta tubulin Ab(#T0023) for 1 hour at 37°C. An AlexaFluor594 conjugated goat anti-rabbit IgG Ab(Red) and an AlexaFluor488 conjugated goat anti-mouse IgG Ab(Green) were used as the secondary antibody.
The nuclear counter stain is DAPI (blue).")

 The protein expression levels of total β-catenin, cytoplasmic β-catenin, nuclear β-catenin and P-β-catenin were detected by western blot.")
 Red fluorescence indicates RFP or PADI3-fused RFP,
and blue indicates nuclei stained with DAPI. The merged image shows both RFP fluorescence and DAPI staining. The scale bar represents
20 μm. (B) Western blot analysis was used to detect truncated PADI3 in the cytoplasm and nucleus. GAPDH was used as the internal control
for cytoplasmic expression, and histone 3 was used as the internal control for nuclear expression. Nu: nucleus, Cy: cytoplasm.")
 and DAPI (blue) in the kidneys from C57BL/6 mice treated with sterilized saline (con), LPS, or LPS plus GSK’872 (GSK) for 24 h. GSK
partially restored the decreased nuclear staining of TFEB in the renal tubular cells of LPS-induced mice. Scale bar = 20 μm. B Cultured PTECs treated
with LPS, LPS plus GSK, LPS plus DMSO, LPS plus RIP3 siRNA, or LPS plus scrambled siRNA (Scramble) for 12 h. The nuclear protein fractions were
immunoblotted for TFEB. Histone was used as the nuclear marker. TFEB expression against Histone was decreased by LPS treatment, which was
restored by GSK or RIP3 siRNA (n = 3). C Immunofluorescence staining for TFEB (green) and DAPI (blue) in cultured PTECs treated as indicated. The
nuclear translocation of TFEB was inhibited by LPS, which was rescued by GSK or RIP3 siRNA. Scale bar = 20 μm. D, E Lysates from cultured PTECs
treated with LPS or DMSO (con) for 12 h were subjected to immunoprecipitation using an anti-RIP3 antibody (D) or anti-TFEB antibody (E), and IgG
antibody followed by immunoblot for TFEB and RIP3. Input proteins were detected with anti-RIP3 and anti-TFEB antibodies. RIP3 interacted with TFEB
in LPS-treated PTECs compared to the control group. F Lysates from cultured PTECs treated with LPS or DMSO (con) for 12 h were subjected to
immunoprecipitation using anti-TFEB antibody and IgG antibody followed by immunoblot for p-RIP3. Input proteins were detected with anti-p-RIP3
and anti-TFEB antibodies. p-RIP3 interacted with TFEB in LPS-treated PTECs compared to the control group. *P < 0.05.")
 schematic diagram, (B) 3D and (C) 2D diagrams of the hydrogen bond interaction between OHC and Keap1; (D) schematic diagram, (E) 3D and (F) 2D diagrams of the hydrogen bond interaction between THC and Keap1; (G) schematic diagram, (H) 3D and (I) 2D diagrams of the hydrogen bond interaction between CUR and Keap1")
 and heme oxygenase‐1 (HO-1) in renal cortical tissues were examined by Western blot analysis. (A, B and C) Nrf2 content in nuclear fraction of renal cortical tissues of rats for 2, 4 and 6wk post-MI by western blot, respectively (mean ± SEM, n = 4 in each group)")
 J774A.1 cells were treated
with lipopolysaccharide (LPS) and pinocembrin for 4 h, and add nigericin (NIG) (10 μM) for 1 h. Images show the protein levels of NLRP3, pro-interleukin 1β (pro-IL-
1β), pro-caspase-1, cleaved-caspase-1, nuclear factor kappa-B (NF-κB). (B-C) RAW264.7 and J774A.1 were treated with LPS and pinocembrin for 4 h. Images show
the expression of NLRP3, pro-IL-1β, NF-κB. Results are shown as mean ± SD (n = 3). #P < 0.05; ##P < 0.01; ###P < 0.001; *P < 0.05; **P < 0.01; ***P < 0.001.")
. The mRNA levels of Nrf-2 in the sham or 5/6 nx groups (B) and the control or adenine groups (C) were detected by RT-PCR. The total protein expression of Nrf-2 under the BSA or AOPPs pretreated (D).Expression of Nrf-2 in cytoplasmic or nuclear (E) were assessed by Western blot. The protein expression of Nrf-2 in the sham or 5/6nx groups (F) and the control or adenine groups (G) were detected by Western blot")
 during LPS stimulation. (A) Bands of the protein related to histone H3 and acetylated histone H3. Quantification of the bands from Western blots.")
 in HepG2 (A) and L-02 cells (B).Cytoplasmic and nuclear protein expression levels of NF-κB were detected by western blotting (C).Protein expression levels were quantified by ImageJ software.")
. DEGs were significantly enriched in the
p53 signaling pathway (b). After treatment with 10-HDAA/LPS for 24 h, the nuclear translocation of p53 was tested after separating nuclear and cytoplasmic
proteins. β-tubulin and Histone-H3 were used as an internal control of cytoplasmic protein and nuclear protein, respectively (c). p-p53 protein levels were tested after
24 h of 10-HDAA and/or LPS treatment using Western blot analysis (d). Cells were transfected with si-NC or si-p53, then the p53 protein level in BV-2 cells was tested
after 48 h (e). Cells were transfected with si-NC or si-p53, 24 h later, cells were treated with 10-HDAA/LPS for 24 h, then IL-1α (f), IL-27 (g), TNF-α (h) and IL-6 (i)
levels in the cell culture medium were tested using ELISA kits. The protein levels of COX-2, iNOS, and TNF-α in BV-2 cells were examined after p53 knockdown (j).
Data are presented as means ± SEM, and group differences were analyzed by one-way ANOVA with post hoc Tukey’s test. ns, nonsignificant, *P < 0.05,
**P < 0.01.")
 The expression of γ-H2AX, TLR2, and p-NF-κB in cells is shown by western blotting (*, p < 0.05, vs. control).
(B) The qRT-PCR results show that TLR2 and NF-κB in Beas-2b cells are blocked by siRNA efficiently. Cells transfected with TLR2 or NF-κB siRNA were treated with
10 μg/mL AFG1 plus 50 ng/mL HMGB1 for 24 h. (C)The expression of γ-H2AX in TLR2-transfected cells after AFG1 plus HMGB1-treatment is shown by western blot
(*, p < 0.05. (D) After AFG1 plus HMGB1-treatment, the expression of γ-H2AX in NF-κB-transfected cells is shown by western blot (*, p < 0.05). (E) After AFG1 plus
HMGB1-treatment, intracellular accumulation of ROS in TLR2- or NF-κB-transfected cells was determined by flow cytometry (*, p < 0.05). (F) After AFG1 plus
HMGB1-treatment, apoptosis in TLR2- or NF-κB-transfected cells was measured by flow cytometry (*, p < 0.05). The data are the mean ± SD of triplicate experiments, representative of three independent experiments (*, p < 0.05).")
 in HepG2 (A) and L-02 cells (B).Cytoplasmic and nuclear protein expression levels of NF-κB were detected by western blotting (C).")
 The negative control or siTP53INP2 were transfected into the BIU87 and EJ cells. The cells were incubated for 48 h, and the changes in the expressions of EMT molecular markers and β-catenin were detected by Western blot. β-actin was used as an internal control. (C, D) The cellular location of active β-catenin and Snail1 in EJ cells after interference in TP53INP2. Scale bar, 50 μm. (E) The abundance of the active forms of β-catenin and Snail1 in nuclear and cytosolic fractions were detected by Western blot. GAPDH and H3 were used as cytosolic or nuclear controls.
Abbreviations: EMT, epithelial-to-mesenchymal transition; GAPDH, glyceraldehyde 3-phosphate dehydrogenase.")
 L02 cells were incubated with various concentrations of crizotinib or sunitinib for 48 h (n = 3). ROS levels were analyzed using a ROS assay kit. (B, C) Total and nuclear Nrf2 levels in L02 cells were measured. (D) C57 mice were gavaged with 100 mg/kg/day crizotinib or 120 mg/kg/day sunitinib for 14 days (n = 6). The levels of GPx, MDA and SOD were measured to assess oxidative stress. (E) The mRNA levels of Nrf2, HO1 and NQO1 were determined by RT-qPCR (n = 8). (F, G) The hepatic protein levels of total and nuclear Nrf2 were determined by western blot analysis (n = 6). *p < 0.05, **p < 0.01 and ***p < 0.001 vs. control group.")
 Western blotting for the total protein.
The level of β-actin protein is taken as loading control. (A1) The relative expression levels of UuMRPS27 normalized to β-actin according to the result of (A). (B)
Western blotting for the nuclear protein. The level of Histone H3 protein is taken as loading control. (B1) The relative contents of nuclear UuMRPS27 normalized to
Histone H3 according to the result of (B). Data are indicated as mean ± SD (n = 3). Asterisk (*) represents significant difference (P < 0.05) between the sulfide
exposure group and control at the same time; Different letters (a-e) among the sulfide exposure groups indicate statistically significant difference (ANOVA, Duncan’s
test; P < 0.05), whilst shared letters indicate no significant difference.")
, **p < 0.01 vs. control.")
 Representative Western blotting images of pIκBα, IκBα, cytoplasmic p65 and nuclear p65 (B–E) The protein expression levels of pIκBα, IκBα, cytoplasmic p65 and nuclear p65 in the NF-κB pathway. Data are expressed as the means ± SD (n = 3). # p < 0.05, ## p < 0.01 vs. control group; *p < 0.05, **p < 0.01 vs. DSS group; & p < 0.05, && p < 0.01 vs. COP group.")
. B, C: Effect of nuclear TTF-1 expression in lung tissue of mice with lung cancer (n=6).")
 and the quantification of the p-NFκB expression in tibias of different groups of rats.
Error bars indicated SEM (n = 6). **P < 0.01 compared with the Mod group. The arrows denoted positive staining. B: Representative western blot images and the
quantification of p-NFκB/NFκB and p-IκB/IκB in the high glucose-induced osteoblasts with treatment with timosaponin BII (0.1 μmol/L, 1 μmol/L, 10 μmol/L) and in
those cells with NAC (25 μmol/L) incubation for 48 h **P < 0.01 compared with the high glucose group. C: Translocation of NFκB by confocal microscopy. Scale bar:
10 μm. D: Representative immunoblotting (upper panel) of protein expression of NFκB in nucleus and cytoplasm and the quantitate data (bottom panel) in osteoblasts
with treatment with timosaponin BII (10 μmol/L). **P < 0.01 compared to the treatment with high glucose alone.")
 Western blot analyses of nucl-Nrf2, cyto-Nrf2, Keap1, nucl-p65, cyto-p65, and p-IκBα (C–F) Intensity of Keap1, cyto-Nrf2, p-IκBα, and cyto-p65 relative to β-actin. Intensity of nucl-Nrf2, nucl-p65 relative to H3. Values represent the means ± SD (n = 3). ## p < 0.01 vs control; * p < 0.05, **p < 0.01 vs TNF-α.")
 Cytosol NF-κB P65; (B) Nuclear NF-κB P65.")

The relative expression level of TLR4. (B) The relative expression level of MyD88. (C) The relative expression level of NF-κB p65. (D) The
relative expression level of nuclear HMGB1. (E) The relative expression level of cytoplasmic HMGB1. Protein samples were acquired from
the ischaemic cortex and assayed by western blot. Nuclear proteins were normalized to the intensity of Histone H3, and cytoplasmic and
total proteins were normalized to the intensity of GAPDH or β-actin. Data were expressed as means ± SD (n = 3). ##P < .01 versus the sham
group; *P < .05 and **P < .01 versus the I/R + PBS group. DPSC-Exos, dental pulp stem cell-derived exosomes; HMGB1, high-mobility group
box 1 protein; I/R, ischaemia/reperfusion; MyD88, myeloid differentiation protein 88; NF-κB, nuclear factor-kappa B; PBS, phosphatebuffered saline; TLR4, toll-like receptor-4")
 and Aβ25-35 on PPARγ expression in PC12 cells. (A) Analysis of PPARγ expression in PC12 cells treated as indicated by
cell labeling with a RBITC-conjugated anti- PPARγ antibody and flow cytometry; (B) Quantification and comparison of PPARγ expression by MFI in (A); All values are
means ± S.E.M. from three independent experiments; **P < 0.01 vs the control; # #P < 0.01 vs the Aβ group, by Student’s T-test. (C) The distribution of PPARγ was
detected in cells by LSCM. The cells were fixed and successively stained with the RBITC-anti-PPARγ antibody (red color) and DAPI (blue color). Cells were treated by
(a) vehicle control (Ctrl), (b) 50 μM GRb1 alone (GRb1), (c) 25 μM Aβ25-35 alone (Aβ), (d) 25 μM GRb1 + 25 μM Aβ25-35 (GRb1-1), and (e) 50 μM GRb1 + 25 μM
Aβ25-35 (GRb1-2), respectively. (D) Detection of cytoplasmic and nuclear expression of PPARγ in PC12 cells by immunoblotting, respectively; (E) Quantitative
analysis of immunoblotting results from (D) by ImageJ software (National Institutes of Health, Bethesda, MA, USA) with α-tublin as cytoplasmic protein loading
control and histone H3 as nuclear protein loading control, respectively. Expression level is shown relative to Ctrl, the value for which was set to 1. All values are
means ± S.E.M. from three independent experiments, **P < 0.01 vs the control; #P < 0.05, ##P < 0.01 and ###P < 0.001 vs the Aβ group, by Student’s T-test.")
. To fit into the manuscript properly, the gel was reasonably trimmed. aa, P < 0.01 vs the control group; b, P < 0.05, bb, P < 0.01 vs the Wnt-1 group. ##, P < 0.01 vs the normal group; *, P < 0.05 and **, P < 0.01 vs the model group")
 The protein expression of Keap1 and nuclear Nrf2 in HepG2 cells (n = 3).")
 Immunohistochemistry staining of p-p65 in the different groups after CLP (Original magnification ×400, scale bar 50 μm). (b) Protein expression levels of p-IκBα, IκBα, p-p65 and nuclear p65 in lung tissue were measured by western blot. Quantitative analysis of the blots was normalized to GAPDH or Histone H3. Results were presented as mean ± standard deviation, n = 6. **P < 0.01 versus Sham group; #P < 0.05 versus CLP group; ##P < 0.01 versus CLP group.")
 Calcitriol decreased myocyte apoptosis and death. Representative photographs (A) and quantitative analysis of TUNEL staining of the cardiomyocytes in mice with the indicated treatments (n = 5). Scale bar represents 50 μm. (C,D) Western blot analysis of protein expressions in the whole-cell lysates (VDR, IL-1β, IL-10, TNF-α, and NF-κB p65) or nuclear lysate (VDR and p-NF-κB p65) in mice with various treatments (n = 5). (E–G) Serum IL-1β, TNF-α, and IL-10 levels in mice with the indicated treatments (n ≥ 8). Data presented here are representative images (A,C) or quantitative analysis (B,D–G), respectively. (Scale bar represents 50 μm. Data represent means ± S.D. * p < 0.05, ** p < 0.01 versus sham group, # p < 0.05, ## p < 0.01 versus MI group; one-way ANOVA with a post hoc test of Tukey’s analysis).")
 Expression of cytokines in serum and liver tissue homogenate. (b) Nuclear proteins from mouse liver tissues were extracted, and p-NF-κB (p65) protein levels in the liver were detected by western blotting. (c) NF-κB relative mRNA levels in the liver. n = 5 per group. Data are presented as the mean ± SD from at least three independent experiments. ∗P < 0.05, ∗∗P < 0.01, ∗∗∗P < 0.005, and ∗∗∗∗P < 0.0001.")
. Immunofluorescent staining was performed on lung sections from CLP mice (n = 3) and analyzed by confocal microscopy. DAPI (blue), NE (Neutrophil elastase, red) and CitH3 (Citrulline histone 3, green) (A,C). The expression of CitH3 in lung tissues of indicated groups of mice. The expression of NE in lung tissues of different groups of mice was shown in (B,D–G). The expression of NETs formation markers, CitH3 and NE, were determined by immunohistochemistry in the lung sections of different groups of mice (F,G). The quantifications of CitH3 (F) and NE (G) expression levels in immunohistochemistry assays (n = 3) (H,I). Western blot was performed to determine the expression of CitH3 in the lung tissue of indicated groups of mice. I showed the quantification of CitH3 expression. N = 4/per group. The values were indicated as mean ± SD. *p < 0.05, **p < 0.01, ***p < 0.001, compared with the CLP group.\"")
 XBJ alleviated the NETs formation in the lungs of septic mice (A–D). Immunofluorescent staining was performed on lung sections from CLP mice (n = 3) and analyzed by confocal microscopy. DAPI (blue), NE (Neutrophil elastase, red) and CitH3 (Citrulline histone 3, green) (A,C). The expression of CitH3 in lung tissues of indicated groups of mice. The expression of NE in lung tissues of different groups of mice was shown in (B,D–G). The expression of NETs formation markers, CitH3 and NE, were determined by immunohistochemistry in the lung sections of different groups of mice (F,G). The quantifications of CitH3 (F) and NE (G) expression levels in immunohistochemistry assays (n = 3) (H,I). Western blot was performed to determine the expression of CitH3 in the lung tissue of indicated groups of mice. I showed the quantification of CitH3 expression. N = 4/per group. The values were indicated as mean ± SD. *p < 0.05, **p < 0.01, ***p < 0.001, compared with the CLP group."" title=""FIGURE 4
XBJ alleviated the NETs formation in the lungs of septic mice (A–D). Immunofluorescent staining was performed on lung sections from CLP mice (n = 3) and analyzed by confocal microscopy. DAPI (blue), NE (Neutrophil elastase, red) and CitH3 (Citrulline histone 3, green) (A,C). The expression of CitH3 in lung tissues of indicated groups of mice. The expression of NE in lung tissues of different groups of mice was shown in (B,D–G). The expression of NETs formation markers, CitH3 and NE, were determined by immunohistochemistry in the lung sections of different groups of mice (F,G). The quantifications of CitH3 (F) and NE (G) expression levels in immunohistochemistry assays (n = 3) (H,I). Western blot was performed to determine the expression of CitH3 in the lung tissue of indicated groups of mice. I showed the quantification of CitH3 expression. N = 4/per group. The values were indicated as mean ± SD. *p < 0.05, **p < 0.01, ***p < 0.001, compared with the CLP group."" alt="Histone H3 Antibody - "FIGURE 4
XBJ alleviated the NETs formation in the lungs of septic mice (A–D).">
XBJ alleviated the NETs formation in the lungs of septic mice (A–D). Immunofluorescent staining was performed on lung sections from CLP mice (n = 3) and analyzed by confocal microscopy. DAPI (blue), NE (Neutrophil elastase, red) and CitH3 (Citrulline histone 3, green) (A,C). The expression of CitH3 in lung tissues of indicated groups of mice. The expression of NE in lung tissues of different groups of mice was shown in (B,D–G). The expression of NETs formation markers, CitH3 and NE, were determined by immunohistochemistry in the lung sections of different groups of mice (F,G). The quantifications of CitH3 (F) and NE (G) expression levels in immunohistochemistry assays (n = 3) (H,I). Western blot was performed to determine the expression of CitH3 in the lung tissue of indicated groups of mice. I showed the quantification of CitH3 expression. N = 4/per group. The values were indicated as mean ± SD. *p < 0.05, **p < 0.01, ***p < 0.001, compared with the CLP group."" title=""FIGURE 4
XBJ alleviated the NETs formation in the lungs of septic mice (A–D). Immunofluorescent staining was performed on lung sections from CLP mice (n = 3) and analyzed by confocal microscopy. DAPI (blue), NE (Neutrophil elastase, red) and CitH3 (Citrulline histone 3, green) (A,C). The expression of CitH3 in lung tissues of indicated groups of mice. The expression of NE in lung tissues of different groups of mice was shown in (B,D–G). The expression of NETs formation markers, CitH3 and NE, were determined by immunohistochemistry in the lung sections of different groups of mice (F,G). The quantifications of CitH3 (F) and NE (G) expression levels in immunohistochemistry assays (n = 3) (H,I). Western blot was performed to determine the expression of CitH3 in the lung tissue of indicated groups of mice. I showed the quantification of CitH3 expression. N = 4/per group. The values were indicated as mean ± SD. *p < 0.05, **p < 0.01, ***p < 0.001, compared with the CLP group."" alt="Histone H3 Antibody - "FIGURE 4
XBJ alleviated the NETs formation in the lungs of septic mice (A–D).">
 ChIP assay. (b) qRT-PCR analysis. (c) Western blotting analysis. A, si-NC; B, si-KMT2D (1); C, si- KMT2D (1).")
 for 4 h, and then treated with Aβ42 oligomers (1 μM) for 24 h, followed by treatment with stigmasterol (20 μM) for 4 h. (A,B) Representative western blot analysis of AMPK signaling. GAPDH immunoreactivity was used as a loading control. (C-E) Representative western blot analysis of NF-κB signaling. The cytosolic and nuclear fractions were prepared and analyzed with total NF-κB p65. α-Tubulin immunoreactivity was used as a loading control in the cytosolic fraction, Histone H3 was used as the loading control in the nuclear fraction. (F-H) Representative western blot analysis of NLRP3 signaling, including NLRP3 and Caspase-1, p20. GAPDH immunoreactivity was used as a loading control (I, J) Concentration of TNFα and IL-1β. Data were presented as the mean ± SEM from three independent experiments. One-way ANOVA with Tukey’s multiple comparison test revealed a difference between groups.")
 RAW264.7 cells were starved with 2% FBS overnight. After treating the cells with cinacalcet (5 μM) for 2 h, TNFα (10 ng/ml) was added for 4 h. Immunofluorescence of P65 was performed (20 μm). White arrow heads indicate the aggregation of P65 in the nuclei. (B) Bone marrow-derived macrophages (BMDMs) were seeded in 10 cm plates. After starvation with 2% FBS overnight, the cells were treated with cinacalcet (0.1, 1, 5 μM) for 2 h followed by TNFα (10 ng/ml) for 45 min. Cytoplasmic and nuclear proteins were extracted for western blotting. (C) Quantification of NE P65 expression standardized to the internal expression of histone H3. (D) Quantification of CE P65 expression standardized to the internal expression of GAPDH. *p < 0.05, **p < 0.01. Data are shown as the mean with standard deviation. Three independent experiments were performed.")
 expression and promotes nuclear translocation of Nrf2 after ICH in vivo and in vitro. (A) Venn analysis of overlapping genes that interact with both WFA and HO-1 in the CTD database and String database. (B) Representative western blot images and (C) quantitative analysis of relative protein level of cytoplasmic Nrf2 and nuclear Nrf2 in the sham, ICH, and WFA (0.1, 1, and 5 μg/kg)-treated groups on day 7 after ICH. (D) Quantification of relative nuclear and cytoplasmic Nrf2 protein levels in the sham, ICH, and WFA (0.1/1/5 μg/kg)-treated groups on day 7 after ICH. (E) Representative western blot images and (F and G) quantification of relative protein expression of cytoplasmic Nrf2, nuclear Nrf2, and ration of nuclear/cytoplasmic Nrf2 in the hemin-induced ICH model with or without WFA treatment. (H) Nrf2 localization was detected by immunofluorescence with an anti-Nrf2 antibody (green fluorescence). Nuclei were stained with DAPI (blue fluorescence). (I) Allocation of green and blue signals in the dual-channel overlay images was analyzed with the ImageJ “Co-localization Finder” plugin, and co-localization of the green and blue signals is shown in white spots. The x-axes indicate the intensities of the green signals from the green channel (Nrf2) and the y-axes from the blue channel (DAPI). For every scatterplot, the intensities are given as the pixel grey values ranging from 0 to 255. Co-localization clusters the pixels from two channels along the diagonal. Pearson’s correlation coefficient (Rr) reflects the intensity distribution relationship between the two channels. Manders overlap coefficient (MOC) reflects the true degree of co-localization of the two channels. The maximal theoretical value for the Rr and MOC is 1.0. Quantification analysis of Nrf2 nuclear location with parameter. (J) Pearson’s correlation coefficient (Rr) and (K) MOC. n = 6 mice per group in B–D; n = 3 per group in E–K. *P < 0.05, **P < 0.01 (one-way analysis of variance followed by Bonferroni correction). Data are presented as the mean ± SEM. HO-1: Heme oxygenase-1; ICH: intracerebral hemorrhage; Nrf2: nuclear factor erythroid-related factor 2; WFA: withaferin A.")
 Naïve CD4+ T cells were cultured under Th17 differentiation conditions with or without MBL protein (5μg/mL). (A) The expression of AhR in CD4+ T cells was determined by FCM analysis. (B) The protein levels of AhR were determined by Western blot. (C) The level of phosphorylated STAT3 was assessed by FCM analysis. (D–E) Naïve CD4+ T cells were pretreated with 10μM Sc144 for 2h, then cultured under Th17 differentiation conditions with or without MBL protein (5μg/mL). (D) The mRNA expression of AhR in CD4+ T cells was determined by RT-qPCR analysis. (E) The protein level of AhR was assessed by Western blot analysis. (F–H) CD4+ T cells were treated with 200nM FICZ for 6h in the presence or absence of 5μg/mL MBL. (F) The fraction of cytoplasm and nucleus protein were separated, and the expression of AhR was determined by Western blot. (G) The mRNA levels of CYP1A1, CYP1A2, and CYP1B1 were detected by RT-qPCR analysis. (H) The mRNA levels of RORγt and IL-17 were detected by RT-qPCR analysis. *p < 0.05, **p < 0.01. The data represent three independent experiments with similar results.")
. Moreover, double immunofluorescence indicated that the TFEB nuclear translocation was mainly displayed in the neurons at the penumbra (E–G). n = 6,")
 expression and 11R-VIVIT could increase the protein expression of p-NFAT2 (Ser172). c Quantification of NFAT2 expression normalized to GAPDH. d Quantification of p-NFAT2 (Ser172) expression normalized to GAPDH. e–f TGFβ treatment increased nuclear NFAT2 expression in HK-2 cells, and 11R-VIVIT inhibited the nuclear localization of NFAT2. g Quantification of NFAT2 expression in the cytoplasmic fraction HK-2 cells; The results were normalized to GAPDH. h Quantification of NFAT2 expression in the nuclear fraction of HK-2 cells; The results were normalized to Histone H3. i The protein expression of α-SMA and fibronectin in HK-2 cells. j–k The quantitative results of α-SMA and fibronectin were normalized to GAPDH. l Apoptosis was examined by flow cytometry. m Quantification of RTEC apoptosis by flow cytometry. n Western blot analysis of the expression of caspase-3 and C‑caspase-3 in HK-2 cells. o The quantitative results of caspase-3 and C‑caspase-3 were normalized to β-actin. p Western blot analysis of the expression of Bax in HK-2 cells. q The quantitative results of Bax were normalized to β-actin. r TUNEL staining of HK-2 cells in the CON, TGFβ and 11R-VIVIT + TGFβ group. s Quantification of TUNEL‑positive tubular epithelial cells. Scale bars = 20 μm. *P")
 The expressions of the CYP2E1, Cytosol Nrf2, HO-1, and nuclear Nrf2 were assessed by Western Blotting (n = 3). β-actin and Histone H3 were used as a loading control. Relative mRNA expression of (b) CYP2E1, (c) HO-1, and (d) NQO1. All data are expressed as mean ± SD (n = 8). #p < 0.05, ##p < 0.01 vs. normal control group; ∗p < 0.05, ∗∗p < 0.01 vs. model group.")
: Detection of MAPK expression and phosphorylation through Western blotting. NCI-H1299 cells were treated with 26 μg/mL of PPD for 48 h. (B): Gray analysis of Western blotting.")
 Representative bands of RANKL, NFATc1, pIκBα, IκBα, cytoplasmic p65, and nuclear p65. (B-F) Relative protein expression levels of RANKL (F-value: 28.501), NFATc1 (F-value: 28.705), pIκBα, IκBα (pIκBα/IκBα, F-value: 10.056), cytoplasmic p65 (F-value:5.692), and nuclear p65 (F-value: 6.893). Data presented as mean ± SD (n = 3), degrees of freedom between groups: 4, degrees of freedom within groups")
 and bar graphs (C-J) of Nrf2, HO-1, TLR4, p-NF-κB p65, NF-κB p65, IL-1β, and IL-6 protein in hippocampus of rats following BZP treatment. Quantification of relative band intensities of total Nrf2, HO-1, nuclear Nrf2 and cytoplasmic Nrf2, and β-actin or histone H3 served as loading control. Values are presented as mean ± SD from 3 separate assays. *p < 0.05, **p < 0.01 vs. sham group; # p < 0.05, ## p < 0.01 vs. 2-VO group; ▲ p < 0.05, ▲▲ p < 0.01 vs. NBP group.")
 H&E staining scores and representative pictures (Scale bar = 100 µm, 50 µm). (B) The TUNEL-positive cells in the lung (Scale bar = 100 µm, 50 µm). Five random fields (200 × ) were evaluated for positive cells. (C) Expression of cytokines in BALF and serum. (D) Nuclear p-p65 protein levels in mouse lung tissues. Use at least three independent experiment to calculate the mean (SD). n = 5. ns, no significance, *p < 0.05, ***p < 0.005, ****p < 0.0001. CLP, cecal ligation puncture; BALF, bronchoalveolar lavage fluid; Dex, Dexmedetomidine; IL, interleukin; TNF, tumor necrosis factor.")
 Western blot and (B) statistical results of western blots of PARP‐1 (n = 5, one‐way ANOVA followed by Bonferroni's post hoc test). (C) Immunofluorescence and (D) statistical analysis of immunofluorescence staining of PARP‐1 in nerve cells from each group (Scale bar = 50 μm, n = 3, one‐way ANOVA followed by Games–Howell post hoc test). (E) Mitochondrial membrane potential was measured with the JC‐1 probe (n = 5, one‐way ANOVA followed by Bonferroni's post hoc test). (F, H, J) Western blot and (G, I, K) statistical results of western blots of Mito‐AIF, Cyto‐AIF, and Nucleo‐AIF (n = 5, one‐way ANOVA followed by Bonferroni's post hoc test). The data represent means ± SD, * means, p")

 was activated in the knee cartilage of 7-day male mice from the maternal obesity group. (A) Plasma leptin level and (B) Lepr mRNA level of knee cartilage 7-day male mice offspring in control versus obesity groups. (C) Correlation between Lepr and cartilage key indicators mRNA expression levels. (D) Representative western blot bands of the expression of leptin receptor in the cell cytoplasm and phosphorylated ERK (p-ERK), phosphorylated p38 (p-38), and phosphorylated c-Jun N-terminal kinase (p-JNK) in the nucleus from knee cartilage of the offspring. (E) Statistical results of the expression of key proteins of MAPK signaling pathway in knee cartilage of the offspring. (A) n = 7, (B) n = 6, (C) n = 6, and (D) n = 6; each band represents the expression of cartilage tissue proteins in two mice. All mice in each group from at least three litters of different mothers. O-WT-P7: offspring 7-day male mice from the control mothers. O-OB-P7: offspring 7-day male mice from obese mothers. Results are expressed as mean ± SD. ns not significant, * p < 0.05, ** p < 0.01, and *** p < 0.001.")
 Western blotting reveals the global lactylation and H3K18la levels of NCAPD3-knockdown H1975 cells and NCAPD3-over-expression HCC827 cells. (B) The protein expression levels of NCAPD3 in H1975 and HCC827 cells with treatment of a series of concentrations of 2-DG or oxamate. (C-D) Proliferation and migration (E) of HCC827 cells with or without NCAPD3 over-expression treated with oxamate (20 mM) were evaluated by CCK-8, colony formation, and transwell assays, respectively. (F-G) Proliferation and migration (H) of H1975 cells with or without NCAPD3 knockdown treated with lactate (5 mM) were evaluated by CCK-8, colony formation, and transwell assays, respectively. ▲P")
 Immunohistochemical staining for NOX4 (300 ×; bar, 50 µm). (B) The relative IOD/area of NOX4. (C) The bands of NOX4, p22phox and p47phox in the human control cerebral artery and IA tissues (Western blot). (D) The relative expressions of NOX4, p22phox and p47phox. (E) The relative mRNA level of NOX4 in the human control cerebral artery and IA tissues (q-PCR). (F) Immunohistochemical staining for TRPC6 (300 ×; bar, 50 µm). (G) The relative IOD/area of TRPC6. (H) The band of TRPC6 in the human control cerebral artery and IA tissues (Western blot). (I) The relative expression of TRPC6. (J) The relative mRNA level of Trpc6 in the human control cerebral artery and IA tissues (q-PCR). (K) The bands of CN and NFATC1 in the human control cerebral artery and IA tissues (Western blot). (L) The relative expressions of CN and NFATC1. (M) The band of NFATC1 nuclear protein in the human control cerebral artery and IA tissues (Western blot). (N) The relative expression of NFATC1 in the nucleus. Data were expressed as mean ± SD, n = 3, * P")
 Effect of Biejiaxiaozheng pills on the Viability of RAW 264.7 Cells. (B) Effect of Biejiaxiaozheng pills on the contents of inflammatory factors in the Supernatant of RAW 264.7 Cells. (C) Effect of Biejiaxiaozheng pills on the Phosphorylation and nuclear translocation of p65 and INOS Protein Expression of RAW 264.7 Cells.")

 Expression and statistical analysis of total hnRNPA2/B1 protein levels. (C, D) Expression and statistical analysis of hnRNPA2/B1 in the nuclei (Nuc) and cytoplasm (Cyto). (E) Immunofluorescence images of hnRNPA2/B1 (red) in the hypoxic hippocampal cells, DAPI (blue), scale bar: 10 μm,2 μm (zoom in image). (F) Quantification analysis of the cytoplasmic distribution of hnRNPA2/B1 using Image J software. Data are presented as mean ± SD (n = 3/group). Student's t-tests was used in B, D, and F. *p")

Control Products
Related Downloads
Protocols
Product Info
*The optimal dilutions should be determined by the end user. For optimal experimental results, antibody reuse is not recommended.
*Tips:
WB: For western blot detection of denatured protein samples. IHC: For immunohistochemical detection of paraffin sections (IHC-p) or frozen sections (IHC-f) of tissue samples. IF/ICC: For immunofluorescence detection of cell samples. ELISA(peptide): For ELISA detection of antigenic peptide.
Cite Format: Affinity Biosciences Cat# AF0863, RRID:AB_2810277.
Fold/Unfold
H3 histone family, member A; H3/A; H31_HUMAN; H3FA; Hist1h3a; HIST1H3B; HIST1H3C; HIST1H3D; HIST1H3E; HIST1H3F; HIST1H3G; HIST1H3H; HIST1H3I; HIST1H3J; histone 1, H3a; Histone cluster 1, H3a; Histone H3.1; Histone H3/a; Histone H3/b; Histone H3/c; Histone H3/d; Histone H3/f; Histone H3/h; Histone H3/i; Histone H3/j; Histone H3/k; Histone H3/l; ;H3.3A; HIST1 cluster, H3E; H3 histone family, member A; H3.1; H3/l; H3F3; H3FF; H3FJ; H3FL; Histone gene cluster 1, H3 histone family, member E; histone H3.1t; Histone H3/o; FLJ92264; H 3; H3; H3 histone family, member B; H3 histone family, member C; H3 histone family, member D; H3 histone family, member F; H3 histone family, member H; H3 histone family, member I; H3 histone family, member J; H3 histone family, member K; H3 histone family, member L; H3 histone family, member T; H3 histone, family 3A; H3/A; H3/b; H3/c; H3/d; h3/f; H3/h; H3/i; H3/j; H3/k; H3/t; H31_HUMAN; H3F1K; H3F3A; H3FA; H3FB; H3FC; H3FD; H3FH; H3FI; H3FK; HIST1 cluster, H3A; HIST1 cluster, H3B; HIST1 cluster, H3C; HIST1 cluster, H3D; HIST1 cluster, H3F; HIST1 cluster, H3G; HIST1 cluster, H3H; HIST1 cluster, H3I; HIST1 cluster, H3J; HIST1H3A; HIST1H3B; HIST1H3C; HIST1H3D; HIST1H3E; HIST1H3F; HIST1H3G; HIST1H3H; HIST1H3I; HIST1H3J; HIST3H3; Histone 1, H3a; Histone 1, H3b; Histone 1, H3c; Histone 1, H3d; Histone 1, H3e; Histone 1, H3f; Histone 1, H3g; Histone 1, H3h; Histone 1, H3i; Histone 3, H3; histone cluster 1 H3 family member a; histone cluster 1 H3 family member b; histone cluster 1 H3 family member c; histone cluster 1 H3 family member d; histone cluster 1 H3 family member e; histone cluster 1 H3 family member f; histone cluster 1 H3 family member g; histone cluster 1 H3 family member h; histone cluster 1 H3 family member i; histone cluster 1 H3 family member j; Histone cluster 1, H3a; Histone cluster 1, H3b; Histone cluster 1, H3c; Histone cluster 1, H3d; Histone cluster 1, H3e; Histone cluster 1, H3f; Histone cluster 1, H3g; Histone cluster 1, H3i; Histone cluster 1, H3j; Histone gene cluster 1, H3 histone family, member A; Histone gene cluster 1, H3 histone family, member B; Histone gene cluster 1, H3 histone family, member C; Histone gene cluster 1, H3 histone family, member D; Histone gene cluster 1, H3 histone family, member F; Histone gene cluster 1, H3 histone family, member G; Histone gene cluster 1, H3 histone family, member H; Histone gene cluster 1, H3 histone family, member I; Histone gene cluster 1, H3 histone family, member J; Histone gene cluster 1, H3A; Histone gene cluster 1, H3B; Histone gene cluster 1, H3C; Histone gene cluster 1, H3D; Histone gene cluster 1, H3E; Histone gene cluster 1, H3F; Histone gene cluster 1, H3G; Histone gene cluster 1, H3H; Histone gene cluster 1, H3I; Histone gene cluster 1, H3J; Histone H 3; Histone H3.1; Histone H3.2; Histone H3.3; Histone H3/a; Histone H3/b; Histone H3/c; Histone H3/d; Histone H3/f; Histone H3/h; Histone H3/i; Histone H3/j; Histone H3/k; Histone H3/l; Histone H3/m; H3 histone family 3A; H3 histone family 3B; H3 histone, family 3B (H3.3B); H3.3; H3.3A; H3.3B; H33_HUMAN; H3F3; H3F3A; H3f3b; Histone H3.3; Histone H3.3Q; Histone H3.A; Histone H3.B; MGC87782; MGC87783;
Immunogens
A synthesized peptide derived from human Histone H3.
- P68431 H31_HUMAN:
- Protein BLAST With
- NCBI/
- ExPASy/
- Uniprot
MARTKQTARKSTGGKAPRKQLATKAARKSAPATGGVKKPHRYRPGTVALREIRRYQKSTELLIRKLPFQRLVREIAQDFKTDLRFQSSAVMALQEACEAYLVGLFEDTNLCAIHAKRVTIMPKDIQLARRIRGERA
- Q71DI3 H32_HUMAN:
- Protein BLAST With
- NCBI/
- ExPASy/
- Uniprot
MARTKQTARKSTGGKAPRKQLATKAARKSAPATGGVKKPHRYRPGTVALREIRRYQKSTELLIRKLPFQRLVREIAQDFKTDLRFQSSAVMALQEASEAYLVGLFEDTNLCAIHAKRVTIMPKDIQLARRIRGERA
- P84243 H33_HUMAN:
- Protein BLAST With
- NCBI/
- ExPASy/
- Uniprot
MARTKQTARKSTGGKAPRKQLATKAARKSAPSTGGVKKPHRYRPGTVALREIRRYQKSTELLIRKLPFQRLVREIAQDFKTDLRFQSAAIGALQEASEAYLVGLFEDTNLCAIHAKRVTIMPKDIQLARRIRGERA
Predictions
Score>80(red) has high confidence and is suggested to be used for WB detection. *The prediction model is mainly based on the alignment of immunogen sequences, the results are for reference only, not as the basis of quality assurance.
High(score>80) Medium(80>score>50) Low(score<50) No confidence
Research Backgrounds
Core component of nucleosome. Nucleosomes wrap and compact DNA into chromatin, limiting DNA accessibility to the cellular machineries which require DNA as a template. Histones thereby play a central role in transcription regulation, DNA repair, DNA replication and chromosomal stability. DNA accessibility is regulated via a complex set of post-translational modifications of histones, also called histone code, and nucleosome remodeling.
Acetylation is generally linked to gene activation. Acetylation on Lys-10 (H3K9ac) impairs methylation at Arg-9 (H3R8me2s). Acetylation on Lys-19 (H3K18ac) and Lys-24 (H3K24ac) favors methylation at Arg-18 (H3R17me). Acetylation at Lys-123 (H3K122ac) by EP300/p300 plays a central role in chromatin structure: localizes at the surface of the histone octamer and stimulates transcription, possibly by promoting nucleosome instability.
Citrullination at Arg-9 (H3R8ci) and/or Arg-18 (H3R17ci) by PADI4 impairs methylation and represses transcription.
Asymmetric dimethylation at Arg-18 (H3R17me2a) by CARM1 is linked to gene activation. Symmetric dimethylation at Arg-9 (H3R8me2s) by PRMT5 is linked to gene repression. Asymmetric dimethylation at Arg-3 (H3R2me2a) by PRMT6 is linked to gene repression and is mutually exclusive with H3 Lys-5 methylation (H3K4me2 and H3K4me3). H3R2me2a is present at the 3' of genes regardless of their transcription state and is enriched on inactive promoters, while it is absent on active promoters.
Methylation at Lys-5 (H3K4me), Lys-37 (H3K36me) and Lys-80 (H3K79me) are linked to gene activation. Methylation at Lys-5 (H3K4me) facilitates subsequent acetylation of H3 and H4. Methylation at Lys-80 (H3K79me) is associated with DNA double-strand break (DSB) responses and is a specific target for TP53BP1. Methylation at Lys-10 (H3K9me) and Lys-28 (H3K27me) are linked to gene repression. Methylation at Lys-10 (H3K9me) is a specific target for HP1 proteins (CBX1, CBX3 and CBX5) and prevents subsequent phosphorylation at Ser-11 (H3S10ph) and acetylation of H3 and H4. Methylation at Lys-5 (H3K4me) and Lys-80 (H3K79me) require preliminary monoubiquitination of H2B at 'Lys-120'. Methylation at Lys-10 (H3K9me) and Lys-28 (H3K27me) are enriched in inactive X chromosome chromatin. Monomethylation at Lys-57 (H3K56me1) by EHMT2/G9A in G1 phase promotes interaction with PCNA and is required for DNA replication.
Phosphorylated at Thr-4 (H3T3ph) by HASPIN during prophase and dephosphorylated during anaphase. Phosphorylation at Ser-11 (H3S10ph) by AURKB is crucial for chromosome condensation and cell-cycle progression during mitosis and meiosis. In addition phosphorylation at Ser-11 (H3S10ph) by RPS6KA4 and RPS6KA5 is important during interphase because it enables the transcription of genes following external stimulation, like mitogens, stress, growth factors or UV irradiation and result in the activation of genes, such as c-fos and c-jun. Phosphorylation at Ser-11 (H3S10ph), which is linked to gene activation, prevents methylation at Lys-10 (H3K9me) but facilitates acetylation of H3 and H4. Phosphorylation at Ser-11 (H3S10ph) by AURKB mediates the dissociation of HP1 proteins (CBX1, CBX3 and CBX5) from heterochromatin. Phosphorylation at Ser-11 (H3S10ph) is also an essential regulatory mechanism for neoplastic cell transformation. Phosphorylated at Ser-29 (H3S28ph) by MAP3K20 isoform 1, RPS6KA5 or AURKB during mitosis or upon ultraviolet B irradiation. Phosphorylation at Thr-7 (H3T6ph) by PRKCB is a specific tag for epigenetic transcriptional activation that prevents demethylation of Lys-5 (H3K4me) by LSD1/KDM1A. At centromeres, specifically phosphorylated at Thr-12 (H3T11ph) from prophase to early anaphase, by DAPK3 and PKN1. Phosphorylation at Thr-12 (H3T11ph) by PKN1 is a specific tag for epigenetic transcriptional activation that promotes demethylation of Lys-10 (H3K9me) by KDM4C/JMJD2C. Phosphorylation at Thr-12 (H3T11ph) by chromatin-associated CHEK1 regulates the transcription of cell cycle regulatory genes by modulating acetylation of Lys-10 (H3K9ac). Phosphorylation at Tyr-42 (H3Y41ph) by JAK2 promotes exclusion of CBX5 (HP1 alpha) from chromatin.
Monoubiquitinated by RAG1 in lymphoid cells, monoubiquitination is required for V(D)J recombination (By similarity). Ubiquitinated by the CUL4-DDB-RBX1 complex in response to ultraviolet irradiation. This may weaken the interaction between histones and DNA and facilitate DNA accessibility to repair proteins.
Lysine deamination at Lys-5 (H3K4all) to form allysine is mediated by LOXL2. Allysine formation by LOXL2 only takes place on H3K4me3 and results in gene repression.
Crotonylation (Kcr) is specifically present in male germ cells and marks testis-specific genes in post-meiotic cells, including X-linked genes that escape sex chromosome inactivation in haploid cells. Crotonylation marks active promoters and enhancers and confers resistance to transcriptional repressors. It is also associated with post-meiotically activated genes on autosomes.
Butyrylation of histones marks active promoters and competes with histone acetylation. It is present during late spermatogenesis.
Succinylation at Lys-80 (H3K79succ) by KAT2A takes place with a maximum frequency around the transcription start sites of genes. It gives a specific tag for epigenetic transcription activation. Desuccinylation at Lys-123 (H3K122succ) by SIRT7 in response to DNA damage promotes chromatin condensation and double-strand breaks (DSBs) repair.
Serine ADP-ribosylation constitutes the primary form of ADP-ribosylation of proteins in response to DNA damage. Serine ADP-ribosylation at Ser-11 (H3S10ADPr) is mutually exclusive with phosphorylation at Ser-11 (H3S10ph) and impairs acetylation at Lys-10 (H3K9ac).
Nucleus. Chromosome.
Belongs to the histone H3 family.
Core component of nucleosome. Nucleosomes wrap and compact DNA into chromatin, limiting DNA accessibility to the cellular machineries which require DNA as a template. Histones thereby play a central role in transcription regulation, DNA repair, DNA replication and chromosomal stability. DNA accessibility is regulated via a complex set of post-translational modifications of histones, also called histone code, and nucleosome remodeling.
Acetylation is generally linked to gene activation. Acetylation on Lys-10 (H3K9ac) impairs methylation at Arg-9 (H3R8me2s). Acetylation on Lys-19 (H3K18ac) and Lys-24 (H3K24ac) favors methylation at Arg-18 (H3R17me). Acetylation at Lys-123 (H3K122ac) by EP300/p300 plays a central role in chromatin structure: localizes at the surface of the histone octamer and stimulates transcription, possibly by promoting nucleosome instability.
Citrullination at Arg-9 (H3R8ci) and/or Arg-18 (H3R17ci) by PADI4 impairs methylation and represses transcription.
Asymmetric dimethylation at Arg-18 (H3R17me2a) by CARM1 is linked to gene activation. Symmetric dimethylation at Arg-9 (H3R8me2s) by PRMT5 is linked to gene repression. Asymmetric dimethylation at Arg-3 (H3R2me2a) by PRMT6 is linked to gene repression and is mutually exclusive with H3 Lys-5 methylation (H3K4me2 and H3K4me3). H3R2me2a is present at the 3' of genes regardless of their transcription state and is enriched on inactive promoters, while it is absent on active promoters.
Methylation at Lys-5 (H3K4me), Lys-37 (H3K36me) and Lys-80 (H3K79me) are linked to gene activation. Methylation at Lys-5 (H3K4me) facilitates subsequent acetylation of H3 and H4. Methylation at Lys-80 (H3K79me) is associated with DNA double-strand break (DSB) responses and is a specific target for TP53BP1. Methylation at Lys-10 (H3K9me) and Lys-28 (H3K27me) are linked to gene repression. Methylation at Lys-10 (H3K9me) is a specific target for HP1 proteins (CBX1, CBX3 and CBX5) and prevents subsequent phosphorylation at Ser-11 (H3S10ph) and acetylation of H3 and H4. Methylation at Lys-5 (H3K4me) and Lys-80 (H3K79me) require preliminary monoubiquitination of H2B at 'Lys-120'. Methylation at Lys-10 (H3K9me) and Lys-28 (H3K27me) are enriched in inactive X chromosome chromatin. Monomethylation at Lys-57 (H3K56me1) by EHMT2/G9A in G1 phase promotes interaction with PCNA and is required for DNA replication.
Phosphorylated at Thr-4 (H3T3ph) by HASPIN during prophase and dephosphorylated during anaphase. Phosphorylation at Ser-11 (H3S10ph) by AURKB is crucial for chromosome condensation and cell-cycle progression during mitosis and meiosis. In addition phosphorylation at Ser-11 (H3S10ph) by RPS6KA4 and RPS6KA5 is important during interphase because it enables the transcription of genes following external stimulation, like mitogens, stress, growth factors or UV irradiation and result in the activation of genes, such as c-fos and c-jun. Phosphorylation at Ser-11 (H3S10ph), which is linked to gene activation, prevents methylation at Lys-10 (H3K9me) but facilitates acetylation of H3 and H4. Phosphorylation at Ser-11 (H3S10ph) by AURKB mediates the dissociation of HP1 proteins (CBX1, CBX3 and CBX5) from heterochromatin. Phosphorylation at Ser-11 (H3S10ph) is also an essential regulatory mechanism for neoplastic cell transformation. Phosphorylated at Ser-29 (H3S28ph) by MAP3K20 isoform 1, RPS6KA5 or AURKB during mitosis or upon ultraviolet B irradiation. Phosphorylation at Thr-7 (H3T6ph) by PRKCB is a specific tag for epigenetic transcriptional activation that prevents demethylation of Lys-5 (H3K4me) by LSD1/KDM1A. At centromeres, specifically phosphorylated at Thr-12 (H3T11ph) from prophase to early anaphase, by DAPK3 and PKN1. Phosphorylation at Thr-12 (H3T11ph) by PKN1 is a specific tag for epigenetic transcriptional activation that promotes demethylation of Lys-10 (H3K9me) by KDM4C/JMJD2C. Phosphorylation at Tyr-42 (H3Y41ph) by JAK2 promotes exclusion of CBX5 (HP1 alpha) from chromatin.
Monoubiquitinated by RAG1 in lymphoid cells, monoubiquitination is required for V(D)J recombination. Ubiquitinated by the CUL4-DDB-RBX1 complex in response to ultraviolet irradiation. This may weaken the interaction between histones and DNA and facilitate DNA accessibility to repair proteins.
Lysine deamination at Lys-5 (H3K4all) to form allysine is mediated by LOXL2. Allysine formation by LOXL2 only takes place on H3K4me3 and results in gene repression.
Crotonylation (Kcr) is specifically present in male germ cells and marks testis-specific genes in post-meiotic cells, including X-linked genes that escape sex chromosome inactivation in haploid cells. Crotonylation marks active promoters and enhancers and confers resistance to transcriptional repressors. It is also associated with post-meiotically activated genes on autosomes.
Butyrylation of histones marks active promoters and competes with histone acetylation. It is present during late spermatogenesis.
Succinylation at Lys-80 (H3K79succ) by KAT2A takes place with a maximum frequency around the transcription start sites of genes. It gives a specific tag for epigenetic transcription activation. Desuccinylation at Lys-123 (H3K122succ) by SIRT7 in response to DNA damage promotes chromatin condensation and double-strand breaks (DSBs) repair.
Serine ADP-ribosylation constitutes the primary form of ADP-ribosylation of proteins in response to DNA damage. Serine ADP-ribosylation at Ser-11 (H3S10ADPr) is mutually exclusive with phosphorylation at Ser-11 (H3S10ph) and impairs acetylation at Lys-10 (H3K9ac).
Nucleus. Chromosome.
Belongs to the histone H3 family.
Variant histone H3 which replaces conventional H3 in a wide range of nucleosomes in active genes. Constitutes the predominant form of histone H3 in non-dividing cells and is incorporated into chromatin independently of DNA synthesis. Deposited at sites of nucleosomal displacement throughout transcribed genes, suggesting that it represents an epigenetic imprint of transcriptionally active chromatin. Nucleosomes wrap and compact DNA into chromatin, limiting DNA accessibility to the cellular machineries which require DNA as a template. Histones thereby play a central role in transcription regulation, DNA repair, DNA replication and chromosomal stability. DNA accessibility is regulated via a complex set of post-translational modifications of histones, also called histone code, and nucleosome remodeling.
Acetylation is generally linked to gene activation. Acetylation on Lys-10 (H3K9ac) impairs methylation at Arg-9 (H3R8me2s). Acetylation on Lys-19 (H3K18ac) and Lys-24 (H3K24ac) favors methylation at Arg-18 (H3R17me). Acetylation at Lys-123 (H3K122ac) by EP300/p300 plays a central role in chromatin structure: localizes at the surface of the histone octamer and stimulates transcription, possibly by promoting nucleosome instability.
Citrullination at Arg-9 (H3R8ci) and/or Arg-18 (H3R17ci) by PADI4 impairs methylation and represses transcription.
Asymmetric dimethylation at Arg-18 (H3R17me2a) by CARM1 is linked to gene activation. Symmetric dimethylation at Arg-9 (H3R8me2s) by PRMT5 is linked to gene repression. Asymmetric dimethylation at Arg-3 (H3R2me2a) by PRMT6 is linked to gene repression and is mutually exclusive with H3 Lys-5 methylation (H3K4me2 and H3K4me3). H3R2me2a is present at the 3' of genes regardless of their transcription state and is enriched on inactive promoters, while it is absent on active promoters.
Specifically enriched in modifications associated with active chromatin such as methylation at Lys-5 (H3K4me), Lys-37 and Lys-80. Methylation at Lys-5 (H3K4me) facilitates subsequent acetylation of H3 and H4. Methylation at Lys-80 (H3K79me) is associated with DNA double-strand break (DSB) responses and is a specific target for TP53BP1. Methylation at Lys-10 (H3K9me) and Lys-28 (H3K27me), which are linked to gene repression, are underrepresented. Methylation at Lys-10 (H3K9me) is a specific target for HP1 proteins (CBX1, CBX3 and CBX5) and prevents subsequent phosphorylation at Ser-11 (H3S10ph) and acetylation of H3 and H4. Methylation at Lys-5 (H3K4me) and Lys-80 (H3K79me) require preliminary monoubiquitination of H2B at 'Lys-120'. Methylation at Lys-10 (H3K9me) and Lys-28 (H3K27me) are enriched in inactive X chromosome chromatin. Monomethylation at Lys-57 (H3K56me1) by EHMT2/G9A in G1 phase promotes interaction with PCNA and is required for DNA replication.
Phosphorylated at Thr-4 (H3T3ph) by HASPIN during prophase and dephosphorylated during anaphase. Phosphorylation at Ser-11 (H3S10ph) by AURKB is crucial for chromosome condensation and cell-cycle progression during mitosis and meiosis. In addition phosphorylation at Ser-11 (H3S10ph) by RPS6KA4 and RPS6KA5 is important during interphase because it enables the transcription of genes following external stimulation, like mitogens, stress, growth factors or UV irradiation and result in the activation of genes, such as c-fos and c-jun. Phosphorylation at Ser-11 (H3S10ph), which is linked to gene activation, prevents methylation at Lys-10 (H3K9me) but facilitates acetylation of H3 and H4. Phosphorylation at Ser-11 (H3S10ph) by AURKB mediates the dissociation of HP1 proteins (CBX1, CBX3 and CBX5) from heterochromatin. Phosphorylation at Ser-11 (H3S10ph) is also an essential regulatory mechanism for neoplastic cell transformation. Phosphorylated at Ser-29 (H3S28ph) by MAP3K20 isoform 1, RPS6KA5 or AURKB during mitosis or upon ultraviolet B irradiation. Phosphorylation at Thr-7 (H3T6ph) by PRKCB is a specific tag for epigenetic transcriptional activation that prevents demethylation of Lys-5 (H3K4me) by LSD1/KDM1A. At centromeres, specifically phosphorylated at Thr-12 (H3T11ph) from prophase to early anaphase, by DAPK3 and PKN1. Phosphorylation at Thr-12 (H3T11ph) by PKN1 is a specific tag for epigenetic transcriptional activation that promotes demethylation of Lys-10 (H3K9me) by KDM4C/JMJD2C. Phosphorylation at Tyr-42 (H3Y41ph) by JAK2 promotes exclusion of CBX5 (HP1 alpha) from chromatin. Phosphorylation on Ser-32 (H3S31ph) is specific to regions bordering centromeres in metaphase chromosomes.
Ubiquitinated. Monoubiquitinated by RAG1 in lymphoid cells, monoubiquitination is required for V(D)J recombination (By similarity).
Lysine deamination at Lys-5 (H3K4all) to form allysine is mediated by LOXL2. Allysine formation by LOXL2 only takes place on H3K4me3 and results in gene repression.
Crotonylation (Kcr) is specifically present in male germ cells and marks testis-specific genes in post-meiotic cells, including X-linked genes that escape sex chromosome inactivation in haploid cells. Crotonylation marks active promoters and enhancers and confers resistance to transcriptional repressors. It is also associated with post-meiotically activated genes on autosomes.
Butyrylation of histones marks active promoters and competes with histone acetylation. It is present during late spermatogenesis.
Succinylation at Lys-80 (H3K79succ) by KAT2A takes place with a maximum frequency around the transcription start sites of genes. It gives a specific tag for epigenetic transcription activation. Desuccinylation at Lys-123 (H3K122succ) by SIRT7 in response to DNA damage promotes chromatin condensation and double-strand breaks (DSBs) repair.
Serine ADP-ribosylation constitutes the primary form of ADP-ribosylation of proteins in response to DNA damage. Serine ADP-ribosylation at Ser-11 (H3S10ADPr) is mutually exclusive with phosphorylation at Ser-11 (H3S10ph) and impairs acetylation at Lys-10 (H3K9ac).
Nucleus. Chromosome.
Specific interaction of trimethylated form at 'Lys-36' (H3.3K36me3) with ZMYND11 is mediated by the encapsulation of Ser-32 residue with a composite pocket formed by the tandem bromo-PWWP domains.
Belongs to the histone H3 family.
Research Fields
· Human Diseases > Substance dependence > Alcoholism.
· Human Diseases > Cancers: Overview > Transcriptional misregulation in cancer.
· Human Diseases > Immune diseases > Systemic lupus erythematosus.
References

Application: WB Species: mouse Sample: B16F10 and 4T1 tumor
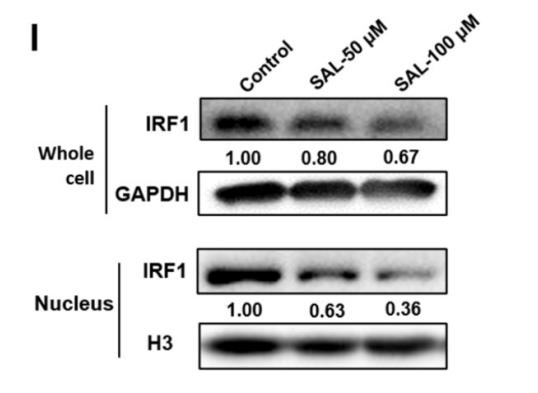
Application: WB Species: human Sample: A375 cells

Application: WB Species: Mouse Sample:

Application: WB Species: Rat Sample: R28 cells

Application: WB Species: Mice Sample: colonic tissues
Restrictive clause
Affinity Biosciences tests all products strictly. Citations are provided as a resource for additional applications that have not been validated by Affinity Biosciences. Please choose the appropriate format for each application and consult Materials and Methods sections for additional details about the use of any product in these publications.
For Research Use Only.
Not for use in diagnostic or therapeutic procedures. Not for resale. Not for distribution without written consent. Affinity Biosciences will not be held responsible for patent infringement or other violations that may occur with the use of our products. Affinity Biosciences, Affinity Biosciences Logo and all other trademarks are the property of Affinity Biosciences LTD.