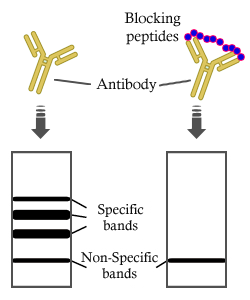
 Antibody at 1/1000 dilution.
5ug/NC membrane strip.
Exposure for 1min with Affinity™ ECL Kit(#KF8003).
Bands result from membrane strip incubation.")
 Antibody.
Lane 1: H2O2 treated MDA-MB-231 cells, blocked with antigen-specific peptides,
Lane 2: H2O2 treated MDA-MB-231 cells,
Lane 3: Heat-shock treated RAW264.7 cells,
Lane 4: UV treated MCF7 cells.")
 Antibody.
Lane 1: HepG2 cells(heat-shock treatment), blocked with antigen-specific peptides.
Lane 2: HepG2 cells(heat-shock treatment).
Lane 3: Raw264.7 cells(serum starvation treatment).")

































| Product: | Acetyl-Histone H2A (Lys5) Antibody |
| Catalog: | AF4354 |
| Description: | Rabbit polyclonal antibody to Acetyl-Histone H2A (Lys5) |
| Application: | WB IHC |
| Reactivity: | Human, Mouse, Rat |
| Mol.Wt.: | 14kDa(Observed); 14kD(Calculated). |
| Uniprot: | P04908 | P0C0S8 | P20671 | Q16777 | Q6FI13 | Q7L7L0 | Q8IUE6 | Q93077 | Q96QV6 | Q99878 |
| RRID: | AB_2844618 |
Control Products
Related Downloads
Protocols
Product Info
*The optimal dilutions should be determined by the end user. For optimal experimental results, antibody reuse is not recommended.
*Tips:
WB: For western blot detection of denatured protein samples. IHC: For immunohistochemical detection of paraffin sections (IHC-p) or frozen sections (IHC-f) of tissue samples. IF/ICC: For immunofluorescence detection of cell samples. ELISA(peptide): For ELISA detection of antigenic peptide.
Cite Format: Affinity Biosciences Cat# AF4354, RRID:AB_2844618.
Fold/Unfold
H2A histone family, member P; H2A.1; H2A.1b; H2A/n; H2A/p; H2A1_HUMAN; H2AF; H2AFC; H2AFD ANDHIST1H2AL; H2AFD; H2AFI; H2AFN ANDHIST1H2AG; H2AFN; H2AFP; HIST1H2AI; HIST1H2AJ; HIST1H2AK; HIST1H2AL; HIST1H2AM; Histone 1, H2ag; Histone cluster 1, H2ag; Histone H2A type 1; Histone H2A.c/d/i/n/p; Histone H2A/p; pH2A/f; H2A histone family member A; H2A histone family member M; H2A.1; H2A.2; H2A/a; H2A/m; H2A1B_HUMAN; H2AFA; H2AFM; HIST1H2AB; HIST1H2AE; Histone 1 H2ab; Histone 1 H2ae; Histone cluster 1 H2ab; Histone cluster 1 H2ae; Histone H2A type 1-B/E; Histone H2A.2; Histone H2A/a; Histone H2A/m; Histone H2AE; H2a 615; H2A; H2A GL101; H2A histone family member A; H2A.1; H2A.2; H2A/a; H2A/m; H2A/O; H2A/q; H2A1B_HUMAN; H2AFA; H2AFE; H2AFL; H2AFM; H2AFO; H2AFQ; HIST1H2AE; HIST1H2AJ; HIST2H2AA; HIST2H2AA3; HIST2H2AB; HIST2H2AC; Histone 1 H2ae; Histone 2 H2aa3; Histone 2 H2ab; Histone 2 H2ac; Histone H2A type 1 B; Histone H2A type 1 C; Histone H2A type 1 E; Histone H2A type 1 J; Histone H2A type 1-B/E; Histone H2A.2; Histone H2A/a; Histone H2A/m; MGC74460; H2a 615; H2A; H2A GL101; H2A histone family member A; H2A.1; H2A.2; H2A/a; H2A/m; H2A/O; H2A/q; H2A1B_HUMAN; H2AFA; H2AFE; H2AFL; H2AFM; H2AFO; H2AFQ; HIST1H2AE; HIST1H2AJ; HIST2H2AA; HIST2H2AA3; HIST2H2AB; HIST2H2AC; Histone 1 H2ae; Histone 2 H2aa3; Histone 2 H2ab; Histone 2 H2ac; Histone H2A type 1 B; Histone H2A type 1 C; Histone H2A type 1 E; Histone H2A type 1 J; Histone H2A type 1-B/E; Histone H2A.2; Histone H2A/a; Histone H2A/m; MGC74460;
Immunogens
A synthesized peptide derived from human Histone H2A around the acetylation site of Lys5.
P04908(H2A1B_HUMAN) >>Visit HPA database.
P0C0S8(H2A1_HUMAN) >>Visit HPA database.
P20671(H2A1D_HUMAN) >>Visit HPA database.
Q16777(H2A2C_HUMAN) >>Visit HPA database.
Q6FI13(H2A2A_HUMAN) >>Visit HPA database.
Q7L7L0(H2A3_HUMAN) >>Visit HPA database.
Q8IUE6(H2A2B_HUMAN) >>Visit HPA database.
Q93077(H2A1C_HUMAN) >>Visit HPA database.
- P04908 H2A1B_HUMAN:
- Protein BLAST With
- NCBI/
- ExPASy/
- Uniprot
MSGRGKQGGKARAKAKTRSSRAGLQFPVGRVHRLLRKGNYSERVGAGAPVYLAAVLEYLTAEILELAGNAARDNKKTRIIPRHLQLAIRNDEELNKLLGRVTIAQGGVLPNIQAVLLPKKTESHHKAKGK
- P0C0S8 H2A1_HUMAN:
- Protein BLAST With
- NCBI/
- ExPASy/
- Uniprot
MSGRGKQGGKARAKAKTRSSRAGLQFPVGRVHRLLRKGNYAERVGAGAPVYLAAVLEYLTAEILELAGNAARDNKKTRIIPRHLQLAIRNDEELNKLLGKVTIAQGGVLPNIQAVLLPKKTESHHKAKGK
- P20671 H2A1D_HUMAN:
- Protein BLAST With
- NCBI/
- ExPASy/
- Uniprot
MSGRGKQGGKARAKAKTRSSRAGLQFPVGRVHRLLRKGNYSERVGAGAPVYLAAVLEYLTAEILELAGNAARDNKKTRIIPRHLQLAIRNDEELNKLLGKVTIAQGGVLPNIQAVLLPKKTESHHKAKGK
- Q16777 H2A2C_HUMAN:
- Protein BLAST With
- NCBI/
- ExPASy/
- Uniprot
MSGRGKQGGKARAKAKSRSSRAGLQFPVGRVHRLLRKGNYAERVGAGAPVYMAAVLEYLTAEILELAGNAARDNKKTRIIPRHLQLAIRNDEELNKLLGKVTIAQGGVLPNIQAVLLPKKTESHKAKSK
- Q6FI13 H2A2A_HUMAN:
- Protein BLAST With
- NCBI/
- ExPASy/
- Uniprot
MSGRGKQGGKARAKAKSRSSRAGLQFPVGRVHRLLRKGNYAERVGAGAPVYMAAVLEYLTAEILELAGNAARDNKKTRIIPRHLQLAIRNDEELNKLLGKVTIAQGGVLPNIQAVLLPKKTESHHKAKGK
- Q7L7L0 H2A3_HUMAN:
- Protein BLAST With
- NCBI/
- ExPASy/
- Uniprot
MSGRGKQGGKARAKAKSRSSRAGLQFPVGRVHRLLRKGNYSERVGAGAPVYLAAVLEYLTAEILELAGNAARDNKKTRIIPRHLQLAIRNDEELNKLLGRVTIAQGGVLPNIQAVLLPKKTESHHKAKGK
- Q8IUE6 H2A2B_HUMAN:
- Protein BLAST With
- NCBI/
- ExPASy/
- Uniprot
MSGRGKQGGKARAKAKSRSSRAGLQFPVGRVHRLLRKGNYAERVGAGAPVYLAAVLEYLTAEILELAGNAARDNKKTRIIPRHLQLAVRNDEELNKLLGGVTIAQGGVLPNIQAVLLPKKTESHKPGKNK
- Q93077 H2A1C_HUMAN:
- Protein BLAST With
- NCBI/
- ExPASy/
- Uniprot
MSGRGKQGGKARAKAKSRSSRAGLQFPVGRVHRLLRKGNYAERVGAGAPVYLAAVLEYLTAEILELAGNAARDNKKTRIIPRHLQLAIRNDEELNKLLGRVTIAQGGVLPNIQAVLLPKKTESHHKAKGK
- Q96QV6 H2A1A_HUMAN:
- Protein BLAST With
- NCBI/
- ExPASy/
- Uniprot
MSGRGKQGGKARAKSKSRSSRAGLQFPVGRIHRLLRKGNYAERIGAGAPVYLAAVLEYLTAEILELAGNASRDNKKTRIIPRHLQLAIRNDEELNKLLGGVTIAQGGVLPNIQAVLLPKKTESHHHKAQSK
- Q99878 H2A1J_HUMAN:
- Protein BLAST With
- NCBI/
- ExPASy/
- Uniprot
MSGRGKQGGKARAKAKTRSSRAGLQFPVGRVHRLLRKGNYAERVGAGAPVYLAAVLEYLTAEILELAGNAARDNKKTRIIPRHLQLAIRNDEELNKLLGKVTIAQGGVLPNIQAVLLPKKTESHHKTK
Research Backgrounds
Core component of nucleosome. Nucleosomes wrap and compact DNA into chromatin, limiting DNA accessibility to the cellular machineries which require DNA as a template. Histones thereby play a central role in transcription regulation, DNA repair, DNA replication and chromosomal stability. DNA accessibility is regulated via a complex set of post-translational modifications of histones, also called histone code, and nucleosome remodeling.
Deiminated on Arg-4 in granulocytes upon calcium entry.
Monoubiquitination of Lys-120 (H2AK119Ub) by RING1, TRIM37 and RNF2/RING2 complex gives a specific tag for epigenetic transcriptional repression and participates in X chromosome inactivation of female mammals. It is involved in the initiation of both imprinted and random X inactivation. Ubiquitinated H2A is enriched in inactive X chromosome chromatin. Ubiquitination of H2A functions downstream of methylation of 'Lys-27' of histone H3 (H3K27me). H2AK119Ub by RNF2/RING2 can also be induced by ultraviolet and may be involved in DNA repair. Monoubiquitination of Lys-120 (H2AK119Ub) by TRIM37 may promote transformation of cells in a number of breast cancers. Following DNA double-strand breaks (DSBs), it is ubiquitinated through 'Lys-63' linkage of ubiquitin moieties by the E2 ligase UBE2N and the E3 ligases RNF8 and RNF168, leading to the recruitment of repair proteins to sites of DNA damage. Ubiquitination at Lys-14 and Lys-16 (H2AK13Ub and H2AK15Ub, respectively) in response to DNA damage is initiated by RNF168 that mediates monoubiquitination at these 2 sites, and 'Lys-63'-linked ubiquitin are then conjugated to monoubiquitin; RNF8 is able to extend 'Lys-63'-linked ubiquitin chains in vitro. Deubiquitinated by USP51 at Lys-14 and Lys-16 (H2AK13Ub and H2AK15Ub, respectively) after damaged DNA is repaired. H2AK119Ub and ionizing radiation-induced 'Lys-63'-linked ubiquitination (H2AK13Ub and H2AK15Ub) are distinct events.
Phosphorylation on Ser-2 (H2AS1ph) is enhanced during mitosis. Phosphorylation on Ser-2 by RPS6KA5/MSK1 directly represses transcription. Acetylation of H3 inhibits Ser-2 phosphorylation by RPS6KA5/MSK1. Phosphorylation at Thr-121 (H2AT120ph) by DCAF1 is present in the regulatory region of many tumor suppresor genes and down-regulates their transcription.
Glutamine methylation at Gln-105 (H2AQ104me) by FBL is specifically dedicated to polymerase I. It is present at 35S ribosomal DNA locus and impairs binding of the FACT complex.
Symmetric dimethylation on Arg-4 by the PRDM1/PRMT5 complex may play a crucial role in the germ-cell lineage.
Crotonylation (Kcr) is specifically present in male germ cells and marks testis-specific genes in post-meiotic cells, including X-linked genes that escape sex chromosome inactivation in haploid cells. Crotonylation marks active promoters and enhancers and confers resistance to transcriptional repressors. It is also associated with post-meiotically activated genes on autosomes.
Nucleus. Chromosome.
Belongs to the histone H2A family.
Core component of nucleosome. Nucleosomes wrap and compact DNA into chromatin, limiting DNA accessibility to the cellular machineries which require DNA as a template. Histones thereby play a central role in transcription regulation, DNA repair, DNA replication and chromosomal stability. DNA accessibility is regulated via a complex set of post-translational modifications of histones, also called histone code, and nucleosome remodeling.
Deiminated on Arg-4 in granulocytes upon calcium entry.
Monoubiquitination of Lys-120 (H2AK119Ub) by RING1, TRIM37 and RNF2/RING2 complex gives a specific tag for epigenetic transcriptional repression and participates in X chromosome inactivation of female mammals. It is involved in the initiation of both imprinted and random X inactivation. Ubiquitinated H2A is enriched in inactive X chromosome chromatin. Ubiquitination of H2A functions downstream of methylation of 'Lys-27' of histone H3 (H3K27me). H2AK119Ub by RNF2/RING2 can also be induced by ultraviolet and may be involved in DNA repair. Monoubiquitination of Lys-120 (H2AK119Ub) by TRIM37 may promote transformation of cells in a number of breast cancers. Following DNA double-strand breaks (DSBs), it is ubiquitinated through 'Lys-63' linkage of ubiquitin moieties by the E2 ligase UBE2N and the E3 ligases RNF8 and RNF168, leading to the recruitment of repair proteins to sites of DNA damage. Ubiquitination at Lys-14 and Lys-16 (H2AK13Ub and H2AK15Ub, respectively) in response to DNA damage is initiated by RNF168 that mediates monoubiquitination at these 2 sites, and 'Lys-63'-linked ubiquitin are then conjugated to monoubiquitin; RNF8 is able to extend 'Lys-63'-linked ubiquitin chains in vitro. Deubiquitinated by USP51 at Lys-14 and Lys-16 (H2AK13Ub and H2AK15Ub, respectively) after damaged DNA is repaired. H2AK119Ub and ionizing radiation-induced 'Lys-63'-linked ubiquitination (H2AK13Ub and H2AK15Ub) are distinct events.
Phosphorylation on Ser-2 (H2AS1ph) is enhanced during mitosis. Phosphorylation on Ser-2 by RPS6KA5/MSK1 directly represses transcription. Acetylation of H3 inhibits Ser-2 phosphorylation by RPS6KA5/MSK1. Phosphorylation at Thr-121 (H2AT120ph) by DCAF1 is present in the regulatory region of many tumor suppresor genes and down-regulates their transcription.
Symmetric dimethylation on Arg-4 by the PRDM1/PRMT5 complex may play a crucial role in the germ-cell lineage.
Glutamine methylation at Gln-105 (H2AQ104me) by FBL is specifically dedicated to polymerase I. It is present at 35S ribosomal DNA locus and impairs binding of the FACT complex.
Crotonylation (Kcr) is specifically present in male germ cells and marks testis-specific genes in post-meiotic cells, including X-linked genes that escape sex chromosome inactivation in haploid cells. Crotonylation marks active promoters and enhancers and confers resistance to transcriptional repressors. It is also associated with post-meiotically activated genes on autosomes.
Nucleus. Chromosome.
Belongs to the histone H2A family.
Core component of nucleosome. Nucleosomes wrap and compact DNA into chromatin, limiting DNA accessibility to the cellular machineries which require DNA as a template. Histones thereby play a central role in transcription regulation, DNA repair, DNA replication and chromosomal stability. DNA accessibility is regulated via a complex set of post-translational modifications of histones, also called histone code, and nucleosome remodeling.
Deiminated on Arg-4 in granulocytes upon calcium entry.
Monoubiquitination of Lys-120 (H2AK119Ub) by RING1, TRIM37 and RNF2/RING2 complex gives a specific tag for epigenetic transcriptional repression and participates in X chromosome inactivation of female mammals. It is involved in the initiation of both imprinted and random X inactivation. Ubiquitinated H2A is enriched in inactive X chromosome chromatin. Ubiquitination of H2A functions downstream of methylation of 'Lys-27' of histone H3 (H3K27me). H2AK119Ub by RNF2/RING2 can also be induced by ultraviolet and may be involved in DNA repair. Monoubiquitination of Lys-120 (H2AK119Ub) by TRIM37 may promote transformation of cells in a number of breast cancers. Following DNA double-strand breaks (DSBs), it is ubiquitinated through 'Lys-63' linkage of ubiquitin moieties by the E2 ligase UBE2N and the E3 ligases RNF8 and RNF168, leading to the recruitment of repair proteins to sites of DNA damage. Ubiquitination at Lys-14 and Lys-16 (H2AK13Ub and H2AK15Ub, respectively) in response to DNA damage is initiated by RNF168 that mediates monoubiquitination at these 2 sites, and 'Lys-63'-linked ubiquitin are then conjugated to monoubiquitin; RNF8 is able to extend 'Lys-63'-linked ubiquitin chains in vitro. Deubiquitinated by USP51 at Lys-14 and Lys-16 (H2AK13Ub and H2AK15Ub, respectively) after damaged DNA is repaired. H2AK119Ub and ionizing radiation-induced 'Lys-63'-linked ubiquitination (H2AK13Ub and H2AK15Ub) are distinct events.
Phosphorylation on Ser-2 (H2AS1ph) is enhanced during mitosis. Phosphorylation on Ser-2 by RPS6KA5/MSK1 directly represses transcription. Acetylation of H3 inhibits Ser-2 phosphorylation by RPS6KA5/MSK1. Phosphorylation at Thr-121 (H2AT120ph) by DCAF1 is present in the regulatory region of many tumor suppresor genes and down-regulates their transcription.
Glutamine methylation at Gln-105 (H2AQ104me) by FBL is specifically dedicated to polymerase I. It is present at 35S ribosomal DNA locus and impairs binding of the FACT complex.
Symmetric dimethylation on Arg-4 by the PRDM1/PRMT5 complex may play a crucial role in the germ-cell lineage.
Crotonylation (Kcr) is specifically present in male germ cells and marks testis-specific genes in post-meiotic cells, including X-linked genes that escape sex chromosome inactivation in haploid cells. Crotonylation marks active promoters and enhancers and confers resistance to transcriptional repressors. It is also associated with post-meiotically activated genes on autosomes.
Nucleus. Chromosome.
Belongs to the histone H2A family.
Core component of nucleosome. Nucleosomes wrap and compact DNA into chromatin, limiting DNA accessibility to the cellular machineries which require DNA as a template. Histones thereby play a central role in transcription regulation, DNA repair, DNA replication and chromosomal stability. DNA accessibility is regulated via a complex set of post-translational modifications of histones, also called histone code, and nucleosome remodeling.
Deiminated on Arg-4 in granulocytes upon calcium entry.
Monoubiquitination of Lys-120 (H2AK119Ub) by RING1, TRIM37 and RNF2/RING2 complex gives a specific tag for epigenetic transcriptional repression and participates in X chromosome inactivation of female mammals. It is involved in the initiation of both imprinted and random X inactivation. Ubiquitinated H2A is enriched in inactive X chromosome chromatin. Ubiquitination of H2A functions downstream of methylation of 'Lys-27' of histone H3 (H3K27me). H2AK119Ub by RNF2/RING2 can also be induced by ultraviolet and may be involved in DNA repair. Monoubiquitination of Lys-120 (H2AK119Ub) by TRIM37 may promote transformation of cells in a number of breast cancers. Following DNA double-strand breaks (DSBs), it is ubiquitinated through 'Lys-63' linkage of ubiquitin moieties by the E2 ligase UBE2N and the E3 ligases RNF8 and RNF168, leading to the recruitment of repair proteins to sites of DNA damage. Ubiquitination at Lys-14 and Lys-16 (H2AK13Ub and H2AK15Ub, respectively) in response to DNA damage is initiated by RNF168 that mediates monoubiquitination at these 2 sites, and 'Lys-63'-linked ubiquitin are then conjugated to monoubiquitin; RNF8 is able to extend 'Lys-63'-linked ubiquitin chains in vitro. Deubiquitinated by USP51 at Lys-14 and Lys-16 (H2AK13Ub and H2AK15Ub, respectively) after damaged DNA is repaired. H2AK119Ub and ionizing radiation-induced 'Lys-63'-linked ubiquitination (H2AK13Ub and H2AK15Ub) are distinct events.
Phosphorylation on Ser-2 (H2AS1ph) is enhanced during mitosis. Phosphorylation on Ser-2 by RPS6KA5/MSK1 directly represses transcription. Acetylation of H3 inhibits Ser-2 phosphorylation by RPS6KA5/MSK1. Phosphorylation at Thr-121 (H2AT120ph) by DCAF1 is present in the regulatory region of many tumor suppresor genes and down-regulates their transcription.
Symmetric dimethylation on Arg-4 by the PRDM1/PRMT5 complex may play a crucial role in the germ-cell lineage.
Glutamine methylation at Gln-105 (H2AQ104me) by FBL is specifically dedicated to polymerase I. It is present at 35S ribosomal DNA locus and impairs binding of the FACT complex.
Crotonylation (Kcr) is specifically present in male germ cells and marks testis-specific genes in post-meiotic cells, including X-linked genes that escape sex chromosome inactivation in haploid cells. Crotonylation marks active promoters and enhancers and confers resistance to transcriptional repressors. It is also associated with post-meiotically activated genes on autosomes.
Nucleus. Chromosome.
Belongs to the histone H2A family.
Core component of nucleosome. Nucleosomes wrap and compact DNA into chromatin, limiting DNA accessibility to the cellular machineries which require DNA as a template. Histones thereby play a central role in transcription regulation, DNA repair, DNA replication and chromosomal stability. DNA accessibility is regulated via a complex set of post-translational modifications of histones, also called histone code, and nucleosome remodeling.
Deiminated on Arg-4 in granulocytes upon calcium entry.
Monoubiquitination of Lys-120 (H2AK119Ub) by RING1, TRIM37 and RNF2/RING2 complex gives a specific tag for epigenetic transcriptional repression and participates in X chromosome inactivation of female mammals. It is involved in the initiation of both imprinted and random X inactivation. Ubiquitinated H2A is enriched in inactive X chromosome chromatin. Ubiquitination of H2A functions downstream of methylation of 'Lys-27' of histone H3 (H3K27me). H2AK119Ub by RNF2/RING2 can also be induced by ultraviolet and may be involved in DNA repair. Monoubiquitination of Lys-120 (H2AK119Ub) by TRIM37 may promote transformation of cells in a number of breast cancers. Following DNA double-strand breaks (DSBs), it is ubiquitinated through 'Lys-63' linkage of ubiquitin moieties by the E2 ligase UBE2N and the E3 ligases RNF8 and RNF168, leading to the recruitment of repair proteins to sites of DNA damage. Ubiquitination at Lys-14 and Lys-16 (H2AK13Ub and H2AK15Ub, respectively) in response to DNA damage is initiated by RNF168 that mediates monoubiquitination at these 2 sites, and 'Lys-63'-linked ubiquitin are then conjugated to monoubiquitin; RNF8 is able to extend 'Lys-63'-linked ubiquitin chains in vitro. Deubiquitinated by USP51 at Lys-14 and Lys-16 (H2AK13Ub and H2AK15Ub, respectively) after damaged DNA is repaired. H2AK119Ub and ionizing radiation-induced 'Lys-63'-linked ubiquitination (H2AK13Ub and H2AK15Ub) are distinct events.
Phosphorylation on Ser-2 (H2AS1ph) is enhanced during mitosis. Phosphorylation on Ser-2 by RPS6KA5/MSK1 directly represses transcription. Acetylation of H3 inhibits Ser-2 phosphorylation by RPS6KA5/MSK1. Phosphorylation at Thr-121 (H2AT120ph) by DCAF1 is present in the regulatory region of many tumor suppresor genes and down-regulates their transcription.
Glutamine methylation at Gln-105 (H2AQ104me) by FBL is specifically dedicated to polymerase I. It is present at 35S ribosomal DNA locus and impairs binding of the FACT complex.
Symmetric dimethylation on Arg-4 by the PRDM1/PRMT5 complex may play a crucial role in the germ-cell lineage.
Crotonylation (Kcr) is specifically present in male germ cells and marks testis-specific genes in post-meiotic cells, including X-linked genes that escape sex chromosome inactivation in haploid cells. Crotonylation marks active promoters and enhancers and confers resistance to transcriptional repressors. It is also associated with post-meiotically activated genes on autosomes.
Nucleus. Chromosome.
Belongs to the histone H2A family.
Core component of nucleosome. Nucleosomes wrap and compact DNA into chromatin, limiting DNA accessibility to the cellular machineries which require DNA as a template. Histones thereby play a central role in transcription regulation, DNA repair, DNA replication and chromosomal stability. DNA accessibility is regulated via a complex set of post-translational modifications of histones, also called histone code, and nucleosome remodeling.
Deiminated on Arg-4 in granulocytes upon calcium entry.
Monoubiquitination of Lys-120 (H2AK119Ub) by RING1, TRIM37 and RNF2/RING2 complex gives a specific tag for epigenetic transcriptional repression and participates in X chromosome inactivation of female mammals. It is involved in the initiation of both imprinted and random X inactivation. Ubiquitinated H2A is enriched in inactive X chromosome chromatin. Ubiquitination of H2A functions downstream of methylation of 'Lys-27' of histone H3 (H3K27me). H2AK119Ub by RNF2/RING2 can also be induced by ultraviolet and may be involved in DNA repair. Monoubiquitination of Lys-120 (H2AK119Ub) by TRIM37 may promote transformation of cells in a number of breast cancers. Following DNA double-strand breaks (DSBs), it is ubiquitinated through 'Lys-63' linkage of ubiquitin moieties by the E2 ligase UBE2N and the E3 ligases RNF8 and RNF168, leading to the recruitment of repair proteins to sites of DNA damage. Ubiquitination at Lys-14 and Lys-16 (H2AK13Ub and H2AK15Ub, respectively) in response to DNA damage is initiated by RNF168 that mediates monoubiquitination at these 2 sites, and 'Lys-63'-linked ubiquitin are then conjugated to monoubiquitin; RNF8 is able to extend 'Lys-63'-linked ubiquitin chains in vitro. Deubiquitinated by USP51 at Lys-14 and Lys-16 (H2AK13Ub and H2AK15Ub, respectively) after damaged DNA is repaired. H2AK119Ub and ionizing radiation-induced 'Lys-63'-linked ubiquitination (H2AK13Ub and H2AK15Ub) are distinct events.
Phosphorylation on Ser-2 (H2AS1ph) is enhanced during mitosis. Phosphorylation on Ser-2 by RPS6KA5/MSK1 directly represses transcription. Acetylation of H3 inhibits Ser-2 phosphorylation by RPS6KA5/MSK1. Phosphorylation at Thr-121 (H2AT120ph) by DCAF1 is present in the regulatory region of many tumor suppresor genes and down-regulates their transcription.
Glutamine methylation at Gln-105 (H2AQ104me) by FBL is specifically dedicated to polymerase I. It is present at 35S ribosomal DNA locus and impairs binding of the FACT complex.
Symmetric dimethylation on Arg-4 by the PRDM1/PRMT5 complex may play a crucial role in the germ-cell lineage.
Crotonylation (Kcr) is specifically present in male germ cells and marks testis-specific genes in post-meiotic cells, including X-linked genes that escape sex chromosome inactivation in haploid cells. Crotonylation marks active promoters and enhancers and confers resistance to transcriptional repressors. It is also associated with post-meiotically activated genes on autosomes.
Nucleus. Chromosome.
Belongs to the histone H2A family.
Core component of nucleosome. Nucleosomes wrap and compact DNA into chromatin, limiting DNA accessibility to the cellular machineries which require DNA as a template. Histones thereby play a central role in transcription regulation, DNA repair, DNA replication and chromosomal stability. DNA accessibility is regulated via a complex set of post-translational modifications of histones, also called histone code, and nucleosome remodeling.
Deiminated on Arg-4 in granulocytes upon calcium entry.
Monoubiquitination of Lys-120 (H2AK119Ub) by RING1, TRIM37 and RNF2/RING2 complex gives a specific tag for epigenetic transcriptional repression and participates in X chromosome inactivation of female mammals. It is involved in the initiation of both imprinted and random X inactivation. Ubiquitinated H2A is enriched in inactive X chromosome chromatin. Ubiquitination of H2A functions downstream of methylation of 'Lys-27' of histone H3 (H3K27me). H2AK119Ub by RNF2/RING2 can also be induced by ultraviolet and may be involved in DNA repair. Monoubiquitination of Lys-120 (H2AK119Ub) by TRIM37 may promote transformation of cells in a number of breast cancers. Following DNA double-strand breaks (DSBs), it is ubiquitinated through 'Lys-63' linkage of ubiquitin moieties by the E2 ligase UBE2N and the E3 ligases RNF8 and RNF168, leading to the recruitment of repair proteins to sites of DNA damage. Ubiquitination at Lys-14 and Lys-16 (H2AK13Ub and H2AK15Ub, respectively) in response to DNA damage is initiated by RNF168 that mediates monoubiquitination at these 2 sites, and 'Lys-63'-linked ubiquitin are then conjugated to monoubiquitin; RNF8 is able to extend 'Lys-63'-linked ubiquitin chains in vitro. Deubiquitinated by USP51 at Lys-14 and Lys-16 (H2AK13Ub and H2AK15Ub, respectively) after damaged DNA is repaired. H2AK119Ub and ionizing radiation-induced 'Lys-63'-linked ubiquitination (H2AK13Ub and H2AK15Ub) are distinct events.
Phosphorylation on Ser-2 (H2AS1ph) is enhanced during mitosis. Phosphorylation on Ser-2 by RPS6KA5/MSK1 directly represses transcription. Acetylation of H3 inhibits Ser-2 phosphorylation by RPS6KA5/MSK1. Phosphorylation at Thr-121 (H2AT120ph) by DCAF1 is present in the regulatory region of many tumor suppresor genes and down-regulates their transcription.
Symmetric dimethylation on Arg-4 by the PRDM1/PRMT5 complex may play a crucial role in the germ-cell lineage.
Glutamine methylation at Gln-105 (H2AQ104me) by FBL is specifically dedicated to polymerase I. It is present at 35S ribosomal DNA locus and impairs binding of the FACT complex.
Crotonylation (Kcr) is specifically present in male germ cells and marks testis-specific genes in post-meiotic cells, including X-linked genes that escape sex chromosome inactivation in haploid cells. Crotonylation marks active promoters and enhancers and confers resistance to transcriptional repressors. It is also associated with post-meiotically activated genes on autosomes.
Nucleus. Chromosome.
Belongs to the histone H2A family.
Core component of nucleosome. Nucleosomes wrap and compact DNA into chromatin, limiting DNA accessibility to the cellular machineries which require DNA as a template. Histones thereby play a central role in transcription regulation, DNA repair, DNA replication and chromosomal stability. DNA accessibility is regulated via a complex set of post-translational modifications of histones, also called histone code, and nucleosome remodeling.
Deiminated on Arg-4 in granulocytes upon calcium entry.
Monoubiquitination of Lys-120 (H2AK119Ub) by RING1, TRIM37 and RNF2/RING2 complex gives a specific tag for epigenetic transcriptional repression and participates in X chromosome inactivation of female mammals. It is involved in the initiation of both imprinted and random X inactivation. Ubiquitinated H2A is enriched in inactive X chromosome chromatin. Ubiquitination of H2A functions downstream of methylation of 'Lys-27' of histone H3 (H3K27me). H2AK119Ub by RNF2/RING2 can also be induced by ultraviolet and may be involved in DNA repair. Monoubiquitination of Lys-120 (H2AK119Ub) by TRIM37 may promote transformation of cells in a number of breast cancers. Following DNA double-strand breaks (DSBs), it is ubiquitinated through 'Lys-63' linkage of ubiquitin moieties by the E2 ligase UBE2N and the E3 ligases RNF8 and RNF168, leading to the recruitment of repair proteins to sites of DNA damage. Ubiquitination at Lys-14 and Lys-16 (H2AK13Ub and H2AK15Ub, respectively) in response to DNA damage is initiated by RNF168 that mediates monoubiquitination at these 2 sites, and 'Lys-63'-linked ubiquitin are then conjugated to monoubiquitin; RNF8 is able to extend 'Lys-63'-linked ubiquitin chains in vitro. Deubiquitinated by USP51 at Lys-14 and Lys-16 (H2AK13Ub and H2AK15Ub, respectively) after damaged DNA is repaired. H2AK119Ub and ionizing radiation-induced 'Lys-63'-linked ubiquitination (H2AK13Ub and H2AK15Ub) are distinct events.
Phosphorylation on Ser-2 (H2AS1ph) is enhanced during mitosis. Phosphorylation on Ser-2 by RPS6KA5/MSK1 directly represses transcription. Acetylation of H3 inhibits Ser-2 phosphorylation by RPS6KA5/MSK1. Phosphorylation at Thr-121 (H2AT120ph) by DCAF1 is present in the regulatory region of many tumor suppresor genes and down-regulates their transcription.
Glutamine methylation at Gln-105 (H2AQ104me) by FBL is specifically dedicated to polymerase I. It is present at 35S ribosomal DNA locus and impairs binding of the FACT complex.
Symmetric dimethylation on Arg-4 by the PRDM1/PRMT5 complex may play a crucial role in the germ-cell lineage.
Crotonylation (Kcr) is specifically present in male germ cells and marks testis-specific genes in post-meiotic cells, including X-linked genes that escape sex chromosome inactivation in haploid cells. Crotonylation marks active promoters and enhancers and confers resistance to transcriptional repressors. It is also associated with post-meiotically activated genes on autosomes.
Nucleus. Chromosome.
Belongs to the histone H2A family.
Core component of nucleosome. Nucleosomes wrap and compact DNA into chromatin, limiting DNA accessibility to the cellular machineries which require DNA as a template. Histones thereby play a central role in transcription regulation, DNA repair, DNA replication and chromosomal stability. DNA accessibility is regulated via a complex set of post-translational modifications of histones, also called histone code, and nucleosome remodeling.
Deiminated on Arg-4 in granulocytes upon calcium entry.
Monoubiquitination of Lys-120 (H2AK119Ub) by RING1, TRIM37 and RNF2/RING2 complex gives a specific tag for epigenetic transcriptional repression and participates in X chromosome inactivation of female mammals. It is involved in the initiation of both imprinted and random X inactivation. Ubiquitinated H2A is enriched in inactive X chromosome chromatin. Ubiquitination of H2A functions downstream of methylation of 'Lys-27' of histone H3 (H3K27me). H2AK119Ub by RNF2/RING2 can also be induced by ultraviolet and may be involved in DNA repair. Monoubiquitination of Lys-120 (H2AK119Ub) by TRIM37 may promote transformation of cells in a number of breast cancers. Following DNA double-strand breaks (DSBs), it is ubiquitinated through 'Lys-63' linkage of ubiquitin moieties by the E2 ligase UBE2N and the E3 ligases RNF8 and RNF168, leading to the recruitment of repair proteins to sites of DNA damage. Ubiquitination at Lys-14 and Lys-16 (H2AK13Ub and H2AK15Ub, respectively) in response to DNA damage is initiated by RNF168 that mediates monoubiquitination at these 2 sites, and 'Lys-63'-linked ubiquitin are then conjugated to monoubiquitin; RNF8 is able to extend 'Lys-63'-linked ubiquitin chains in vitro. Deubiquitinated by USP51 at Lys-14 and Lys-16 (H2AK13Ub and H2AK15Ub, respectively) after damaged DNA is repaired. H2AK119Ub and ionizing radiation-induced 'Lys-63'-linked ubiquitination (H2AK13Ub and H2AK15Ub) are distinct events.
Phosphorylation on Ser-2 (H2AS1ph) is enhanced during mitosis. Phosphorylation on Ser-2 by RPS6KA5/MSK1 directly represses transcription. Acetylation of H3 inhibits Ser-2 phosphorylation by RPS6KA5/MSK1. Phosphorylation at Thr-121 (H2AT120ph) by DCAF1 is present in the regulatory region of many tumor suppresor genes and down-regulates their transcription.
Glutamine methylation at Gln-105 (H2AQ104me) by FBL is specifically dedicated to polymerase I. It is present at 35S ribosomal DNA locus and impairs binding of the FACT complex.
Symmetric dimethylation on Arg-4 by the PRDM1/PRMT5 complex may play a crucial role in the germ-cell lineage.
Crotonylation (Kcr) is specifically present in male germ cells and marks testis-specific genes in post-meiotic cells, including X-linked genes that escape sex chromosome inactivation in haploid cells. Crotonylation marks active promoters and enhancers and confers resistance to transcriptional repressors. It is also associated with post-meiotically activated genes on autosomes.
Nucleus. Chromosome.
Belongs to the histone H2A family.
Core component of nucleosome. Nucleosomes wrap and compact DNA into chromatin, limiting DNA accessibility to the cellular machineries which require DNA as a template. Histones thereby play a central role in transcription regulation, DNA repair, DNA replication and chromosomal stability. DNA accessibility is regulated via a complex set of post-translational modifications of histones, also called histone code, and nucleosome remodeling.
Deiminated on Arg-4 in granulocytes upon calcium entry.
Monoubiquitination of Lys-120 (H2AK119Ub) by RING1, TRIM37 and RNF2/RING2 complex gives a specific tag for epigenetic transcriptional repression and participates in X chromosome inactivation of female mammals. It is involved in the initiation of both imprinted and random X inactivation. Ubiquitinated H2A is enriched in inactive X chromosome chromatin. Ubiquitination of H2A functions downstream of methylation of 'Lys-27' of histone H3 (H3K27me). H2AK119Ub by RNF2/RING2 can also be induced by ultraviolet and may be involved in DNA repair. Monoubiquitination of Lys-120 (H2AK119Ub) by TRIM37 may promote transformation of cells in a number of breast cancers. Following DNA double-strand breaks (DSBs), it is ubiquitinated through 'Lys-63' linkage of ubiquitin moieties by the E2 ligase UBE2N and the E3 ligases RNF8 and RNF168, leading to the recruitment of repair proteins to sites of DNA damage. Ubiquitination at Lys-14 and Lys-16 (H2AK13Ub and H2AK15Ub, respectively) in response to DNA damage is initiated by RNF168 that mediates monoubiquitination at these 2 sites, and 'Lys-63'-linked ubiquitin are then conjugated to monoubiquitin; RNF8 is able to extend 'Lys-63'-linked ubiquitin chains in vitro. Deubiquitinated by USP51 at Lys-14 and Lys-16 (H2AK13Ub and H2AK15Ub, respectively) after damaged DNA is repaired. H2AK119Ub and ionizing radiation-induced 'Lys-63'-linked ubiquitination (H2AK13Ub and H2AK15Ub) are distinct events.
Phosphorylation on Ser-2 (H2AS1ph) is enhanced during mitosis. Phosphorylation on Ser-2 by RPS6KA5/MSK1 directly represses transcription. Acetylation of H3 inhibits Ser-2 phosphorylation by RPS6KA5/MSK1. Phosphorylation at Thr-121 (H2AT120ph) by DCAF1 is present in the regulatory region of many tumor suppresor genes and down-regulates their transcription.
Glutamine methylation at Gln-105 (H2AQ104me) by FBL is specifically dedicated to polymerase I. It is present at 35S ribosomal DNA locus and impairs binding of the FACT complex.
Symmetric dimethylation on Arg-4 by the PRDM1/PRMT5 complex may play a crucial role in the germ-cell lineage.
Crotonylation (Kcr) is specifically present in male germ cells and marks testis-specific genes in post-meiotic cells, including X-linked genes that escape sex chromosome inactivation in haploid cells. Crotonylation marks active promoters and enhancers and confers resistance to transcriptional repressors. It is also associated with post-meiotically activated genes on autosomes.
Nucleus. Chromosome.
Belongs to the histone H2A family.
Research Fields
· Cellular Processes > Cell growth and death > Necroptosis. (View pathway)
· Human Diseases > Substance dependence > Alcoholism.
· Human Diseases > Immune diseases > Systemic lupus erythematosus.
Restrictive clause
Affinity Biosciences tests all products strictly. Citations are provided as a resource for additional applications that have not been validated by Affinity Biosciences. Please choose the appropriate format for each application and consult Materials and Methods sections for additional details about the use of any product in these publications.
For Research Use Only.
Not for use in diagnostic or therapeutic procedures. Not for resale. Not for distribution without written consent. Affinity Biosciences will not be held responsible for patent infringement or other violations that may occur with the use of our products. Affinity Biosciences, Affinity Biosciences Logo and all other trademarks are the property of Affinity Biosciences LTD.