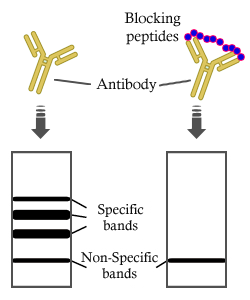
, using Phospho-Rpb1 CTD (Ser2/Ser5) Antibody. The lane on the left was treated with blocking peptide.")
, using Phospho-Rpb1 CTD (Ser2/Ser5) Antibody at 1/1000 dilution.
5ug/NC membrane strip.
Exposure for 5s with Affinity™ ECL Kit(#KF8003).
Bands result from membrane strip incubation.")



 by IF/ICC. The samples were fixed with PFA and permeabilized in 0.1% Triton X-100,then blocked in 10% serum for 45 minutes at 25°C. Samples were then incubated with primary Ab(AF7272 1:200) and mouse anti-beta tubulin Ab(T0023 1:200) for 1 hour at 37°C. An AlexaFluor594 conjugated goat anti-rabbit IgG(H+L) Ab(Red) and an AlexaFluor488 conjugated goat anti-mouse IgG(H+L) Ab(Green) were used as the secondary antibody.
The nuclear counter stain is DAPI(blue).")

| Product: | Phospho-Rpb1 CTD (Ser2/Ser5) Antibody |
| Catalog: | AF7272 |
| Description: | Rabbit polyclonal antibody to Phospho-Rpb1 CTD (Ser2/Ser5) |
| Application: | WB IHC IF/ICC |
| Reactivity: | Human, Mouse, Rat, Monkey |
| Prediction: | Pig, Zebrafish, Bovine, Horse, Sheep, Dog, Xenopus |
| Mol.Wt.: | 270kDa(Observed); 217kD(Calculated). |
| Uniprot: | P24928 |
| RRID: | AB_2843712 |
Control Products
Related Downloads
Protocols
Product Info
*The optimal dilutions should be determined by the end user. For optimal experimental results, antibody reuse is not recommended.
*Tips:
WB: For western blot detection of denatured protein samples. IHC: For immunohistochemical detection of paraffin sections (IHC-p) or frozen sections (IHC-f) of tissue samples. IF/ICC: For immunofluorescence detection of cell samples. ELISA(peptide): For ELISA detection of antigenic peptide.
Cite Format: Affinity Biosciences Cat# AF7272, RRID:AB_2843712.
Fold/Unfold
DNA directed RNA polymerase II subunit RPB1; hRPB220; hsRPB1; POLR2; POLR2A; POLRA; polymerase (RNA) II (DNA directed) polypeptide A, 220kDa; RNA polymerase II subunit 1; RNA polymerase II subunit B1; RP02; RPB220; RPBh1; RpIILS; RPO21; RPOL2; SUA8;
Immunogens
A synthesized peptide derived from human Rpb1 CTD around the phosphorylation site of Ser2/5.
- P24928 RPB1_HUMAN:
- Protein BLAST With
- NCBI/
- ExPASy/
- Uniprot
MHGGGPPSGDSACPLRTIKRVQFGVLSPDELKRMSVTEGGIKYPETTEGGRPKLGGLMDPRQGVIERTGRCQTCAGNMTECPGHFGHIELAKPVFHVGFLVKTMKVLRCVCFFCSKLLVDSNNPKIKDILAKSKGQPKKRLTHVYDLCKGKNICEGGEEMDNKFGVEQPEGDEDLTKEKGHGGCGRYQPRIRRSGLELYAEWKHVNEDSQEKKILLSPERVHEIFKRISDEECFVLGMEPRYARPEWMIVTVLPVPPLSVRPAVVMQGSARNQDDLTHKLADIVKINNQLRRNEQNGAAAHVIAEDVKLLQFHVATMVDNELPGLPRAMQKSGRPLKSLKQRLKGKEGRVRGNLMGKRVDFSARTVITPDPNLSIDQVGVPRSIAANMTFAEIVTPFNIDRLQELVRRGNSQYPGAKYIIRDNGDRIDLRFHPKPSDLHLQTGYKVERHMCDGDIVIFNRQPTLHKMSMMGHRVRILPWSTFRLNLSVTTPYNADFDGDEMNLHLPQSLETRAEIQELAMVPRMIVTPQSNRPVMGIVQDTLTAVRKFTKRDVFLERGEVMNLLMFLSTWDGKVPQPAILKPRPLWTGKQIFSLIIPGHINCIRTHSTHPDDEDSGPYKHISPGDTKVVVENGELIMGILCKKSLGTSAGSLVHISYLEMGHDITRLFYSNIQTVINNWLLIEGHTIGIGDSIADSKTYQDIQNTIKKAKQDVIEVIEKAHNNELEPTPGNTLRQTFENQVNRILNDARDKTGSSAQKSLSEYNNFKSMVVSGAKGSKINISQVIAVVGQQNVEGKRIPFGFKHRTLPHFIKDDYGPESRGFVENSYLAGLTPTEFFFHAMGGREGLIDTAVKTAETGYIQRRLIKSMESVMVKYDATVRNSINQVVQLRYGEDGLAGESVEFQNLATLKPSNKAFEKKFRFDYTNERALRRTLQEDLVKDVLSNAHIQNELEREFERMREDREVLRVIFPTGDSKVVLPCNLLRMIWNAQKIFHINPRLPSDLHPIKVVEGVKELSKKLVIVNGDDPLSRQAQENATLLFNIHLRSTLCSRRMAEEFRLSGEAFDWLLGEIESKFNQAIAHPGEMVGALAAQSLGEPATQMTLNTFHYAGVSAKNVTLGVPRLKELINISKKPKTPSLTVFLLGQSARDAERAKDILCRLEHTTLRKVTANTAIYYDPNPQSTVVAEDQEWVNVYYEMPDFDVARISPWLLRVELDRKHMTDRKLTMEQIAEKINAGFGDDLNCIFNDDNAEKLVLRIRIMNSDENKMQEEEEVVDKMDDDVFLRCIESNMLTDMTLQGIEQISKVYMHLPQTDNKKKIIITEDGEFKALQEWILETDGVSLMRVLSEKDVDPVRTTSNDIVEIFTVLGIEAVRKALERELYHVISFDGSYVNYRHLALLCDTMTCRGHLMAITRHGVNRQDTGPLMKCSFEETVDVLMEAAAHGESDPMKGVSENIMLGQLAPAGTGCFDLLLDAEKCKYGMEIPTNIPGLGAAGPTGMFFGSAPSPMGGISPAMTPWNQGATPAYGAWSPSVGSGMTPGAAGFSPSAASDASGFSPGYSPAWSPTPGSPGSPGPSSPYIPSPGGAMSPSYSPTSPAYEPRSPGGYTPQSPSYSPTSPSYSPTSPSYSPTSPNYSPTSPSYSPTSPSYSPTSPSYSPTSPSYSPTSPSYSPTSPSYSPTSPSYSPTSPSYSPTSPSYSPTSPSYSPTSPSYSPTSPSYSPTSPSYSPTSPSYSPTSPSYSPTSPNYSPTSPNYTPTSPSYSPTSPSYSPTSPNYTPTSPNYSPTSPSYSPTSPSYSPTSPSYSPSSPRYTPQSPTYTPSSPSYSPSSPSYSPASPKYTPTSPSYSPSSPEYTPTSPKYSPTSPKYSPTSPKYSPTSPTYSPTTPKYSPTSPTYSPTSPVYTPTSPKYSPTSPTYSPTSPKYSPTSPTYSPTSPKGSTYSPTSPGYSPTSPTYSLTSPAISPDDSDEEN
Predictions
Score>80(red) has high confidence and is suggested to be used for WB detection. *The prediction model is mainly based on the alignment of immunogen sequences, the results are for reference only, not as the basis of quality assurance.
High(score>80) Medium(80>score>50) Low(score<50) No confidence
Research Backgrounds
DNA-dependent RNA polymerase catalyzes the transcription of DNA into RNA using the four ribonucleoside triphosphates as substrates. Largest and catalytic component of RNA polymerase II which synthesizes mRNA precursors and many functional non-coding RNAs. Forms the polymerase active center together with the second largest subunit. Pol II is the central component of the basal RNA polymerase II transcription machinery. It is composed of mobile elements that move relative to each other. RPB1 is part of the core element with the central large cleft, the clamp element that moves to open and close the cleft and the jaws that are thought to grab the incoming DNA template. At the start of transcription, a single-stranded DNA template strand of the promoter is positioned within the central active site cleft of Pol II. A bridging helix emanates from RPB1 and crosses the cleft near the catalytic site and is thought to promote translocation of Pol II by acting as a ratchet that moves the RNA-DNA hybrid through the active site by switching from straight to bent conformations at each step of nucleotide addition. During transcription elongation, Pol II moves on the template as the transcript elongates. Elongation is influenced by the phosphorylation status of the C-terminal domain (CTD) of Pol II largest subunit (RPB1), which serves as a platform for assembly of factors that regulate transcription initiation, elongation, termination and mRNA processing. Regulation of gene expression levels depends on the balance between methylation and acetylation levels of tha CTD-lysines (By similarity). Initiation or early elongation steps of transcription of growth-factors-induced immediate early genes are regulated by the acetylation status of the CTD. Methylation and dimethylation have a repressive effect on target genes expression (By similarity).
(Microbial infection) Acts as an RNA-dependent RNA polymerase when associated with small delta antigen of Hepatitis delta virus, acting both as a replicate and transcriptase for the viral RNA circular genome.
The tandem heptapeptide repeats in the C-terminal domain (CTD) can be highly phosphorylated. The phosphorylation activates Pol II. Phosphorylation occurs mainly at residues 'Ser-2' and 'Ser-5' of the heptapeptide repeat and is mediated, at least, by CDK7 and CDK9. CDK7 phosphorylation of POLR2A associated with DNA promotes transcription initiation by triggering dissociation from DNA. Phosphorylation also takes place at 'Ser-7' of the heptapeptide repeat, which is required for efficient transcription of snRNA genes and processing of the transcripts. The phosphorylation state is believed to result from the balanced action of site-specific CTD kinases and phosphatases, and a 'CTD code' that specifies the position of Pol II within the transcription cycle has been proposed. Dephosphorylated by the protein phosphatase CTDSP1.
Among tandem heptapeptide repeats of the C-terminal domain (CTD) some do not match the Y-S-P-T-S-P-S consensus, the seventh serine residue 'Ser-7' being replaced by a lysine. 'Lys-7' in these non-consensus heptapeptide repeats can be alternatively acetylated, methylated and dimethylated. EP300 is one of the enzyme able to acetylate 'Lys-7'. Acetylation at 'Lys-7' of non-consensus heptapeptide repeats is associated with 'Ser-2' phosphorylation and active transcription. Regulates initiation or early elongation steps of transcription specially for inducible genes.
Methylated at Arg-1810 prior to transcription initiation when the CTD is hypophosphorylated, phosphorylation at Ser-1805 and Ser-1808 preventing this methylation. Symmetrically or asymmetrically dimethylated at Arg-1810 by PRMT5 and CARM1 respectively. Symmetric or asymmetric dimethylation modulates interactions with CTD-binding proteins like SMN1/SMN2 and TDRD3. SMN1/SMN2 interacts preferentially with the symmetrically dimethylated form while TDRD3 interacts with the asymmetric form. Through the recruitment of SMN1/SMN2, symmetric dimethylation is required for resolving RNA-DNA hybrids created by RNA polymerase II, that form R-loop in transcription terminal regions, an important step in proper transcription termination. CTD dimethylation may also facilitate the expression of select RNAs. Among tandem heptapeptide repeats of the C-terminal domain (CTD) some do not match the Y-S-P-T-S-P-S consensus, the seventh serine residue 'Ser-7' being replaced by a lysine. 'Lys-7' in these non-consensus heptapeptide repeats can be alternatively acetylated, methylated, dimethylated and trimethylated. Methylation occurs in the earliest transcription stages and precedes or is concomitant to 'Ser-5' and 'Ser-7' phosphorylation. Dimethylation and trimehtylation at 'Lys-7' of non-consensus heptapeptide repeats are exclusively associated with phosphorylated CTD.
Ubiquitinated by WWP2 leading to proteasomal degradation (By similarity). Following UV treatment, the elongating form of RNA polymerase II (RNA pol IIo) is ubiquitinated on UV damage sites without leading to degradation: ubiquitination is facilitated by KIAA1530/UVSSA and promotes RNA pol IIo backtracking to allow access to the nucleotide excision repair machinery.
Nucleus. Cytoplasm.
Note: Hypophosphorylated form is mainly found in the cytoplasm, while the hyperphosphorylated and active form is nuclear.
The C-terminal domain (CTD) serves as a platform for assembly of factors that regulate transcription initiation, elongation, termination and mRNA processing.
Belongs to the RNA polymerase beta' chain family.
Research Fields
· Genetic Information Processing > Transcription > RNA polymerase.
· Human Diseases > Neurodegenerative diseases > Huntington's disease.
· Human Diseases > Infectious diseases: Viral > Herpes simplex infection.
· Human Diseases > Infectious diseases: Viral > Epstein-Barr virus infection.
· Metabolism > Nucleotide metabolism > Purine metabolism.
· Metabolism > Nucleotide metabolism > Pyrimidine metabolism.
· Metabolism > Global and overview maps > Metabolic pathways.
Restrictive clause
Affinity Biosciences tests all products strictly. Citations are provided as a resource for additional applications that have not been validated by Affinity Biosciences. Please choose the appropriate format for each application and consult Materials and Methods sections for additional details about the use of any product in these publications.
For Research Use Only.
Not for use in diagnostic or therapeutic procedures. Not for resale. Not for distribution without written consent. Affinity Biosciences will not be held responsible for patent infringement or other violations that may occur with the use of our products. Affinity Biosciences, Affinity Biosciences Logo and all other trademarks are the property of Affinity Biosciences LTD.